ОБЩАЯ ХАРАКТЕРИСТИКА ЖЕНСКОЙ ПОЛОВОЙ СИСТЕМЫ
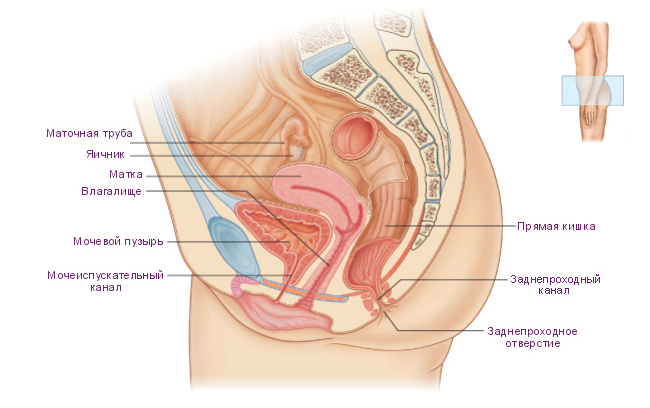
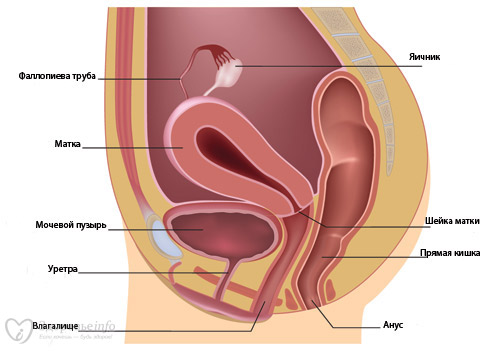
Женская половая система состоит из внутренних и наружных половых органов. К внутренним половым органам, расположенным в полости малого таза, относятся яичники, маточные трубы, матка и влагалище. Наружные половые органы представлены клитором, преддверием влагалища, большими и малыми половыми губами.
Женские половые органы претерпевают регулярные циклические изменения, называемые менструальным циклом. Первый менструальный цикл, менархе, наступает в период от 9 до 14 лет (в среднем в 12,7 лет) и указывает на начало репродуктивного цикла жизни женщины. В возрасте от 45 до 55 лет (в среднем в 51,4 год) менструальные циклы становятся нерегулярными и постепенно исчезают. Эти изменения репродуктивной функции обозначаются как менопауза или климакс.
В период жизни женщины между менархе и наступлением менопаузы примерно каждые 28 дней происходят циклические изменения структуры и активности большинства половых органов, в особенности яичников (овариальный цикл) и матки (менструальный цикл). Синхронизация этих циклов, критичная для нормальной репродуктивной функции женщины, контролируется, главным образом, гипофизарными гонадотропинами: фолликулостимулирующим гормоном (ФСГ) и лютеинизирующим гормоном (ЛГ). Эти гормоны напрямую воздействуют на яичники, модулируя как рост и развитие фолликулов, так и продукцию гормонов яичником. Гормоны яичника (эстрогены и прогестерон) контролируют циклические изменения в эндометрии матки и влияют на продукцию гипофизарного гонадотропина посредством отрицательной обратной связи.
В период менопаузы угасают репродуктивная (образование яйцеклетки) и эндокринная (продукция гормонов) функции яичников. Другие органы (например, влагалище и молочные железы) так же подвергаются изменениям различной степени выраженности.
Молочные железы рассматриваются совместно с репродуктивной системой, так как их развитие и функциональное состояние напрямую зависят от гормональной активности женского организма. Молочные железы подвергаются гистологическим изменениям при менструальном цикле, беременности под действием гормонов гипофиза и яичника. По этой же причине будут рассмотрены избранные аспекты раннего эмбрионального развития: предимплатационное развитие эмбриона, имплантация, а также формирование и функции плаценты.
ЯИЧНИКИ
Яичники – это половые железы (гонады) женского организма, являются парными органами, имеют миндалевидную форму и средние размеры 3х1,5х1 см, расположены в полости малого таза. Снаружи яичники покрыты герминативным эпителием, который, несмотря на своё название, не участвует в образовании ооцитов. Герминативный эпителий представлен однослойным кубическим эпителием, образующимся из мезотелия брюшины. Кнутри от герминативного эпителия располагается плотная соединительная ткань – белочная оболочка.
Каждый яичник состоит из периферически расположенного коркового и центрально лежащего мозгового вещества. В корковом веществе находятся фолликулы яичника, погружённые в соединительную ткань (строму). Мозговое вещество представлено рыхлой соединительной тканью, богатой сетью кровеносных, лимфатических сосудов и нервов. Граница между корковым и мозговым веществом чётко не визуализируется.
Главными функциями яичника являются продукция гамет (гаметогенез) и синтез стероидных гормонов. В женском организме процесс образования гамет называется оогенез (овогенез), развивающиеся гаметы – ооциты, зрелые гаметы – яйцеклетки.
Яичники продуцируют две группы стероидных гормонов: эстрогены и прогестерон. Эти гормоны играют важную роль в менструальном цикле, подготавливая матку к имплантации плодного яйца.
Под действием эстрогенов (эстрон, эстриол и эстрадиол) происходит рост и созревание внутренних и наружных половых органов, а также формирование вторичных половых признаков в пубертатный период. Эстрогены влияют на молочные железы, стимулируя развитие протоковых и стромальных структур, и накопление жировой ткани.
Прогестерон подготавливает внутренние половые органы, в особенности матку, к беременности, при этом происходят секреторные изменения эндометрия (описаны в разделе Менструальный цикл). Кроме того, прогестероны подготавливают молочные железы к лактации, запуская лобулярную пролиферацию.
Фолликулы яичника
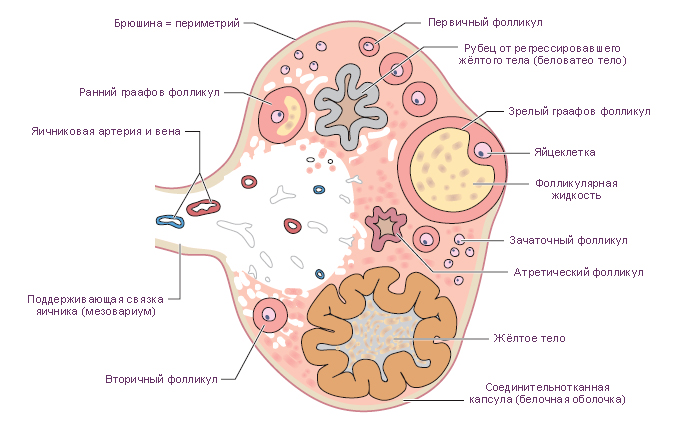
Фолликулы яичника содержат в себе по одному ооциту и создают микроокружение для его развития. Фолликулы бывают разных размеров, и распределены в строме коркового вещества. Размер фолликула отражает стадию его развития. Ранние стадии оогенеза проходят в фетальный период развития, во время которого путём митотических делений массивно увеличивается число оогоний. Развитие ооцитов, находящихся в яичниках при рождении, останавливается на стадии первого мейотического деления. Во время пубертатного периода группа фолликулов проходит этапы роста и созревания. Циклический паттерн созревания фолликулов и овуляции (овариальный цикл) синхронизируется с менструальным циклом.
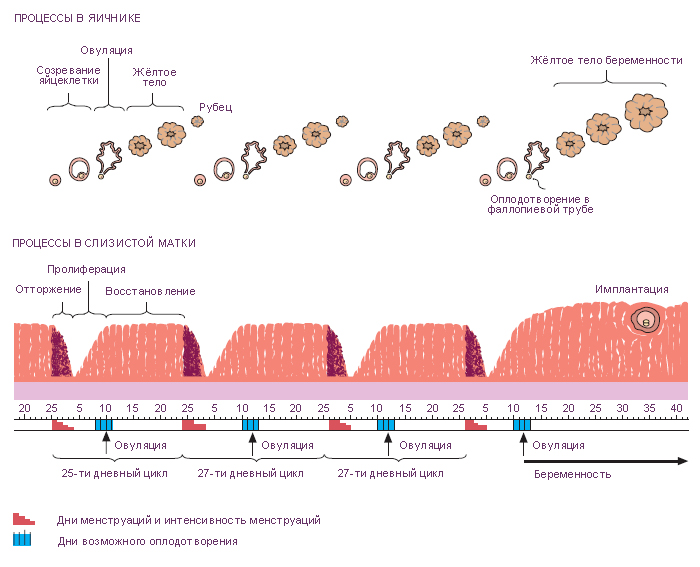
Овариальный цикл – это изменения структуры и активности яичников в репродуктивный период жизни женщины, происходящие каждые 28 дней (в случае отсутствия беременности). Овариальный цикл регулируют гипофизарные гонадотропины – ФСГ и ЛГ. Первая половина овариального цикла – фолликулярная (под влиянием ФСГ происходит развитие части примордиальных фолликулов), вторая половина – лютеиновая (под влиянием ЛГ из клеток овулировавшего фолликула формируется временная эндокринная железа – жёлтое тело). Овуляция приходится примерно на середину цикла. Овариальный цикл сопровождается характерными изменениями содержания половых гормонов в крови.
В нормальных условиях во время каждого овариального цикла активируются примерно 20 фолликулов, из которых только один становится доминантным и созревает. Созревание и выход нескольких ооцитов во время овуляции может привести к образованию многочисленных зигот. В репродуктивный период жизни женщины полного созревания достигают примерно 450 яйцеклеток. Большинство первичных ооцитов, заложенных с рождения (около 1 млн), не завершают полного развития и погибают. Ооциты, оставшиеся после менопаузы, дегенерируют в течение нескольких лет.
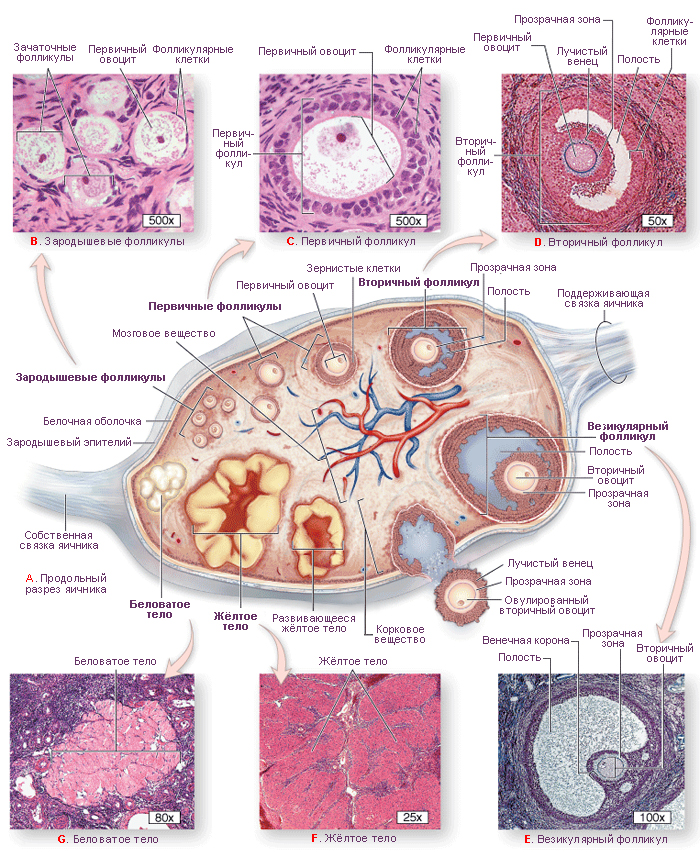
Этапы развития фолликулов (фолликулогенез). Гистологически в зависимости от стадии развития могут быть идентифицированы три типа овариальных фолликулов:
примордиальные фолликулы;
растущие фолликулы, которые подразделяются на первичные, вторичные и третичные (антральные) фолликулы; зрелые или Граафовы фолликулы.
В яичнике можно обнаружить фолликулы на всех стадиях развития.
Примордиальные фолликулы – это самая ранняя стадия развития фолликулов. Они впервые появляются в яичниках на третьем месяце внутриутробного развития. До пубертатного периода в яичниках обнаруживаются только неактивные примордиальные фолликулы, они же составляют большинство и в последующем. В зрелом яичнике примордиальные фолликулы лежат в строме коркового вещества сразу под белочной оболочкой.
Каждый примордиальный фолликул содержит первичный ооцит
(большинство на стадии диплотены профазы первого деления мейоза), который окружён одним слоем плоских фолликулярных клеток. Снаружи от фолликулярных клеток располагается базальная мембрана. На данном этапе размер ооцита составляет 30 мкм, ядро ооцита большое и смещено на периферию.
Растущие фолликулы. Рост фолликулов стимулируется ФСГ.
а. Первичные фолликулы содержат первичный ооцит, окружённый одним слоем кубических фолликулярных клеток. На этой стадии ооцит увеличивается в размерах (до 80 мкм) и начинает секретировать специфичные белки, выделяющиеся из клетки. Между ооцитом и прилежащими фолликулярными клетками образуется блестящая оболочка (zona pellucida, ZP), богатая гликопротеинами ZP-1, ZP-2, ZP-3. Самый важный гликопротеин ZP-3 является рецептором, связывающим сперматозоид и запускающим акросомную реакцию; предполагается, что ZP-2 является вторичным рецептором сперматозоидов, а функциональная роль ZP-1 ещё не была охарактеризована.
б. Вторичные фолликулы. После быстрых митотических делений из одного слоя фолликулярных клеток образуется многослойный кубический эпителий. Базальная мембрана смещается и располагается между наружными слоями фолликулярных клеток и соединительнотканной стромой. Между фолликулярными клетками формируются многочисленные щелевые контакты для обмена питательными веществами и информационными макромолекулами. Во время пролиферации фолликулярных клеток, стромальные клетки, окружающие фолликул, формируют новую оболочку, называемую текой фолликула. Эта оболочка располагается кнаружи от базальной мембраны.
в. Третичные (антральные) фолликулы характеризуются наличием заполненной жидкостью полости. Из фолликулярных клеток формируется гранулёза (фолликулярные клетки с этого момента называются гранулёзными клетками). Гранулёзные клетки начинают секретировать фолликулярную жидкость, в результате чего между ними образуются заполненные жидкостью мелкие полости, которые постепенно сливаются в одну большую полость – антрум. Первичный ооцит смещается к одному из полюсов фолликула, увеличивается до 125-150 мкм в диаметре и перестаёт расти, фолликул же достигает 10 мм и более в диаметре.
Тека фолликула разделяется на два слоя:
Внутренняя тека – внутренний слой кубических секреторных клеток с хорошо развитой гладкой эндоплазматической сетью. Эти клетки содержат большое количество рецепторов ЛГ. В ответ на стимуляцию ЛГ, эндокринные клетки теки синтезируют и секретируют андрогены, являющиеся предшественниками эстрогенов. Часть андрогенов транспортируются к гранулёзным клеткам, которые в ответ на стимуляцию ФСГ под действием ароматазы превращают андрогены в эстрогены. Кроме секреторных клеток, внутренняя тека содержит фибробласты, пучки волокон коллагена и многочисленные капилляры, типичные для эндокринных органов.
Наружная тека – наружный слой, состоящий из фибробластов, гладкомышечных клеток и пучков коллагеновых волокон.
г. Зрелые (Граафовы) фолликулы отличаются от третичных фолликулов своими крупными размерами (10 мм и более в диаметре). Эта стадия предшествует овуляции. Примерно за 24-36 ч до овуляции происходит пик выброса ЛГ аденогипофизом, это вызывает завершение первого мейотического деления первичным ооцитом, что в итоге приводит к образованию вторичного ооцита и первого полярного тельца.
По периметру фолликула гранулёза имеет примерно одинаковую толщину, за исключением зоны, ассоциированной с ооцитом. В этом месте гранулёзные клетки образуют утолщение, вдающееся в полость фолликула и называемое яйценосный бугорок (cumulus oophorus) – «пьедестал» ооцита.
Гранулёзные клетки яйценосного бугорка, окружающие ооцит, становятся лучистым венцом (corona radiata).
3. Атретические фолликулы. Хотя при рождении в яичниках имеется около 1 млн фолликулов, только 400-450 достигают зрелости. Более 99 % становятся атретическими, т.е. они дегенерируют путём аутолиза на разных стадиях развития. При атрезии первичных фолликулов на их месте образуется пространство, которое заполняется стромой и заживает без образования рубцов. Остатки аутолиза первичных и вторичных фолликулов удаляются макрофагами и замещаются стромальными клетками с образованием коллагенового рубца. Рубец постепенно ремоделируется в нормальную строму. Некоторые клетки теки из атретических фолликулов могут сохранять способность синтезировать стероиды, в частности андрогены.
Овуляция
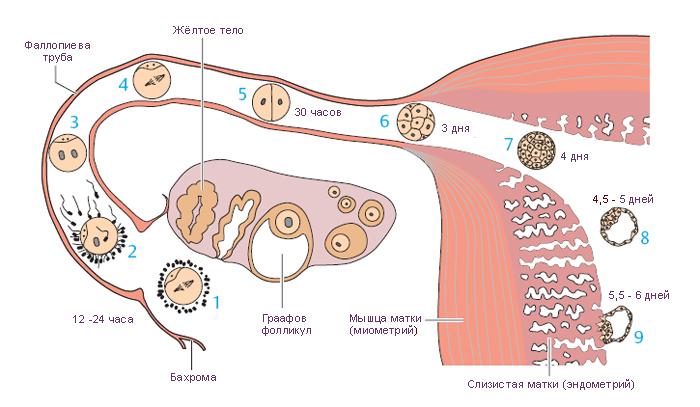
Овуляция – это гормонально-зависимый процесс высвобождения вторичного ооцита из Граафова фолликула. Происходит овуляция в середину менструального цикла (на 14-й день при 28-дневном цикле). Этому предшествует быстрый рост продукции гипофизарного ЛГ с увеличением количества жидкости и давления в антруме фолликула. Благодаря коллагеназной активности, строма между фолликулом и поверхностью яичника истончается, становится ишемической (плохо кровоснабжаемой), указывая на место неизбежного разрыва (фолликулярная стигма). Вторичный ооцит, окружённый лучистым венцом и гранулёзными клетками яйценосного бугорка, высвобождается из разорвавшегося фолликула и захватывается бахромками маточной трубы. После овуляции ооцит сохраняет жизнеспособность в течение примерно 24 часов. Если оплодотворение не происходит в этот период, вторичный ооцит дегенерирует в маточной трубе.
Происхождение и созревание ооцитов
Эндодерма желточного мешка дает начало примордиальным половым клеткам (мужским и женским), которые мигрируют в половые валики на задней стенке брюшной полости (эмбриональные гонады). После миграции клетки дифференцируются и индуцируют дифференцировку яичников (яичек у мужчин). Половые клетки окружены уплощёнными фолликулярными клетками примордиальных фолликулов; они вступают в первое деление мейоза и останавливаются на стадии диплотены профазы на 12-50 лет. На этом этапе они называются первичными ооцитами (сравнимы с первичными сперматоцитами). Первое деление мейоза заканчивается непосредственно перед овуляцией в зрелом фолликуле, при этом происходит равномерное деление хроматина и неравное деление цитоплазмы между образовавшимися дочерними клетками. Первая клетка с большим количеством цитоплазмы называется вторичным ооцитом; вторая клетка с меньшим количеством цитоплазмы называется первым полярным тельцем.
После своего формирования, но до начала овуляции, вторичный ооцит вступает во второе мейотическое деление, которое останавливается в метафазе до наступления оплодотворения. В момент оплодотворения заканчивается второе деление мейоза и образуется яйцеклетка и второе полярное тельце. У человека первое полярное тельце сохраняется более 20 часов и не делится, оплодотворённая яйцеклетка (зигота) содержит два полярных тельца (диплоидное первое и гаплоидное второе полярные тельца). Полярные тельца в дальнейшем подвергаются апоптозу.
Жёлтое тело
Жёлтое тело – временная эндокринная железа, которая образуется из остатков фолликула после овуляции. С его появлением начинается лютеиновая фаза овариального цикла. Во время разрыва фолликула выстилка из гранулёзных клеток и клеток теки образует складки; кровотечение из капилляров внутренней теки в просвет фолликула останавливается с образованием центрального тромба (геморрагическое тело). В ответ на это, клетки стромы проникают в оставшуюся полость фолликула.
Клетки гранулёзного слоя и внутренней теки дифференцируются в гранулёзные лютеиновые клетки и текальные лютеиновые клетки, соответственно. Эти лютеиновые клетки изменяются морфологически, они увеличиваются в размерах, заполняются липидными каплями и начинают секретировать стероидные гормоны:
гранулёзные лютеиновые клетки крупные, бледные при окраске, расположены центрально и синтезируют прогестерон;
текальные лютеиновые клетки меньше в размерах, тёмные при окраске, лежат по периферии и синтезируют эстрогены.
Жёлтое тело дегенерирует примерно через 14 дней, в случае, если оплодотворение не произошло и называется жёлтым телом менструации.
Если оплодотворение происходит, жёлтое тело увеличивается в размерах и функционирует в течение 6 месяцев; после этого оно постепенно уменьшается, но сохраняется до конца беременности. Размеры жёлтого тела в ранние сроки беременности достигают 2-3 см, оно заполняет собой почти весь яичник и называется жёлтое тело беременности. Высокие уровни прогестерона, продуцируемого лютеиновыми клетками, блокируют новые циклы созревания фолликулов.
Белое тело. Этот рубец из плотной соединительной ткани, который замещает дегенерировавшее жёлтое тело. Как и атретические фолликулы, белое тело постепенно разрушается макрофагами.
Гормональная регуляция овариального цикла
Гипофизарный ФСГ стимулирует рост фолликулов в первую фазу менструального цикла. ЛГ стимулирует синтез андрогенов клетками внутренней теки, андрогены в гранулёзных клетках под действием ФСГ трансформируются в эстрогены. Растущие фолликулы продуцируют эстрогены, высокие концентрации которых в середину цикла влияют на продукцию ФСГ по механизму отрицательной обратной связи. В это время резко возрастает концентрация ЛГ, который отвечает за конечное созревание фолликулов, стимулирует овуляцию, а также контролирует формирование и функционирование жёлтого тела. Жёлтое тело синтезирует как эстрогены, так и прогестерон. Прогестерон ингибирует продукцию ЛГ, что приводит к дегенерации жёлтого тела примерно на 14-й день, если оплодотворение не произошло. Если же яйцеклетка была оплодотворена и имплантирована в матку, хорионический гонадотропин человека (ХГЧ), вырабатываемый трофобластом и сходный по строению и функцией с ЛГ, поддерживает жёлтое тело в отсутствии ЛГ.
МАТОЧНЫЕ ТРУБЫ
Маточные трубы (Фаллопиевы трубы, яйцеводы) – это парные трубчатые мышечные органы, проксимальный просвет которых сообщается с полостью матки, а дистальный конец каждой трубы открывается в брюшную полость около яичника.
Маточные трубы приближаются к яичникам перед овуляцией и захватывают овулировавшую яйцеклетку. По маточной трубе яйцеклетка перемещается в полость матки, в ней созданы благоприятные условия для оплодотворения, и транспорта зиготы и морулы, в матку.
Длина каждой маточной трубы составляет 10-12 см, она подразделяется на четыре сегмента:
Маточная (интрамуральная) часть проникает в стенку матки. Этот участок, длиной около 1 см, содержит немногочисленные складки слизистой оболочки, а миометрий участвует в образовании мышечного слоя.
Перешеек – узкий сегмент трубы, прилежащий к стенке матки, содержит небольшое количество складок слизистой оболочки.
Следующая за перешейком широкая часть – ампула, содержит большие ветвящиеся складки слизистой оболочки и является обычным местом оплодотворения. Длина этого сегмента составляет 2/3 общей длины маточной трубы.
Воронка является дистальным сегментом маточной трубы, просвет которого открывается в брюшную полость и заканчивается около яичника пальцевидными разрастаниями – бахромками (фимбрии).
Строение стенки. Стенка маточной трубы состоит из трёх слоев:
Слизистая оболочка – внутренняя выстилка маточной трубы, складки которой выступают в просвет органа. Продольные складки наиболее крупные и многочисленные в ампуле, уменьшаются в размерах и количестве по направлению к матке. Слизистая оболочка состоит из однослойного цилиндрического эпителия и собственной пластики. Эпителий содержит три типа клеток: реснитчатые, секреторные и «крючковидные» (peg cells). Относительное число каждого клеточного типа зависит не столько от региона трубы, сколько от фазы менструального цикла. Эстрогены стимулируют образование реснитчатых клеток, прогестероны – секреторных клеток. Реснитчатые клетки доминируют в середине цикла, и их число резко уменьшается и минимально в течение менструации. Биение ресничек на поверхности цилиндрических клеток осуществляется в направлении матки и способствует транспорту яйцеклетки. Секреторные клетки обычно более узкие, чем реснитчатые, имеют гиперхромное ядро, и вакуолизированный апикальный полюс, нарастают в количестве по направлению к маточной части трубы. Пленка из слизи, которую они синтезируют, продвигается по направлению к матке с помощью ресничек, помогая транспорту яйцеклетки и препятствуя проникновению бактерий в брюшную полость. «Крючковидные» клетки – это самые редкие эпителиальные клетки маточной трубы (<10% от общего количества клеток), которые имеют гиперхромное ядро и небольшое количество цитоплазмы. Реснички на апикальном полюсе клеток отсутствуют. Функция их неясна. Подслизистая оболочка отсутствует. Собственная пластинка слизистой оболочки, образующая основу складок, построена из рыхлой соединительной ткани, богатой кровеносными сосудами.
Мышечная оболочка состоит из двух слоев гладких мышц: внутреннего, относительно толстого, циркулярного и наружного, более тонкого, продольного. Граница между этими слоями часто видна не отчётливо. Волнообразные сокращения этой оболочки продвигают яйцеклетку к матке.
Снаружи маточные трубы покрыты серозной оболочкой – частью широкой связки матки, имеется брыжейка.
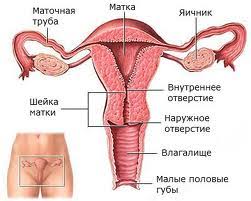
МАТКА
Матка – полый мышечный орган грушевидной формы, расположенный в полости малого таза, служит местом имплантации и развития зародыша. Анатомически в матке выделяют три области. Тело представлено большой средней частью органа конусовидной формы. Дно – это часть тела матки выше впадения маточных труб. Узкое продолжение матки, открывающееся во влагалище носит название шейки матки.
Строение стенки матки
Стенка матки состоит из трёх слоёв:
Эндометрий (слизистая оболочка матки) представлен однослойным цилиндрическим эпителием с подлежащей собственной пластинкой. Толщина эндометрия варьирует от 1 до 6 мм в разные фазы менструального цикла. В состав эпителия входят реснитчатые и секреторные клетки. Поверхностный эпителий проникает в подлежащую собственную пластинку, формируя простые трубчатые маточные железы. Так же, как и в стенке маточной трубы, подслизистая оболочка отсутствует.
Функционально и структурно эндометрий подразделяется на два слоя. Функциональный слой является временным, в ответ на действие гормонов яичника этот слой утолщается и в последующем отторгается при менструации. В зависимости от плотности собственной пластинки этот слой может быть подразделён на компактную зону, лежащую ближе к просвету, и глубокую губчатую зону. Базальный слой – это тонкий постоянный слой, который лежит глубже функционального, сохраняется при менструации и служит источником регенерации функционального слоя. Он содержит базальные отделы маточных желез. Эпителиальные клетки, выстилающие железы базального слоя, делятся митозом и заселяют поверхность собственной пластинки, обнажившуюся при менструации.
Эндометрий получает двойное кровоснабжение. В толще миометрия парные маточные артерии ветвятся с образованием дуговых артерий. Ветви дуговых артерий, радиальные артерии, проникают в базальный слой эндометрия и дают начало двум видам артерий: прямые артерии кровоснабжают базальный слой, спиральные артерии – функциональный слой эндометрия. Двойное кровоснабжение играет важную роль при циклических изменениях функционального слоя, когда спиральные артерии отторгаются вместе с эпителием, а прямые остаются.
Миометрий. Самая толстая мышечная оболочка матки состоит из трёх плохо дифференцируемых друг от друга слоёв гладких мышц.
Средний слой содержит крупные кровеносные и лимфатические сосуды(дуговые артерии и венозные сплетения) и называется сосудистый слой. Пучки ГМК этого слоя расположены циркулярно. Внутренний и наружный слои мышц ориентированы параллельно длинной оси матки.
Во время беременности миометрий интенсивно растёт за счёт гипертрофии и гиперплазии. Пик секреции окситоцина индуцирует мощные сокращения миометрия для изгнания плода во время родов, и три слоя гладких мышц работают как функциональный синцитий.
Периметрий (серозная оболочка) и адвентиция. Дно и большая часть тела покрыты периметрием, нижние отделы тела и надвлагалищная часть шейки матки — адвентицией.
Менструальный цикл
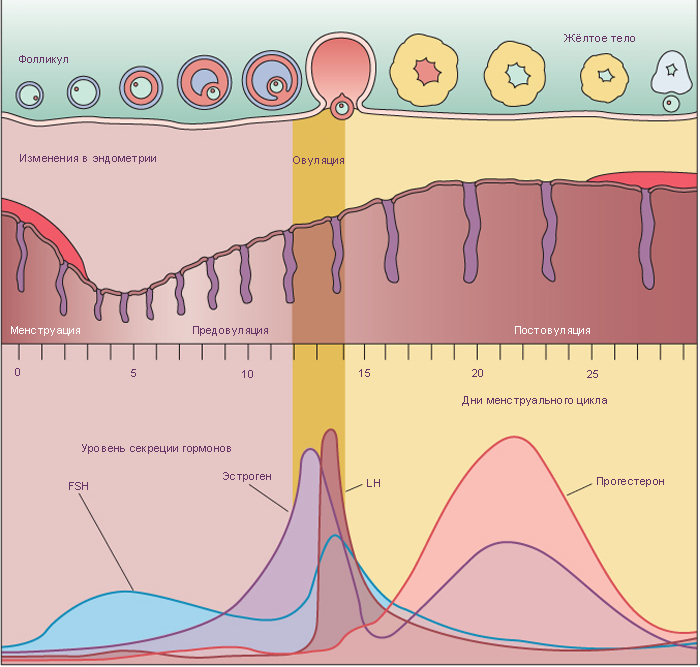
Менструальный цикл – это циклические изменения эндометрия, происходящие каждые 28 дней и подготавливающие матку к имплантации и последующему развитию зародыша, контролируемые овариальными гормонами: эстрогенами и прогестероном. Продукция гормонов яичника, в свою очередь, контролируется гормонами гипофиза: ФСГ и ЛГ, которые регулируют рост фолликулов, овуляцию, образование и дегенерацию жёлтого тела (т.е. овариальный цикл). Менструальный цикл подразделяется на три фазы в зависимости от структурных и функциональных изменений эндометрия: пролиферативная, секреторная и менструальная фазы. В конце каждого цикла происходит частичная деструкция и отторжение эндометрия, сопровождающееся кровотечением из артерий слизистой оболочки. Выделение отделившегося эпителия и крови из влагалища обычно занимает 3-5 дней и называется менструация или менструальное кровотечение. Началом менструального цикла считается первый день менструации. Смена фаз происходит плавно.
В конце менструальной фазы эндометрий представлен только базальным слоем, состоящим из тонкой пластинки соединительной ткани, толщиной примерно 1 мм, в которую погружены базальные отделы маточных желёз и начальные отделы спиральных артерий. Эпителий желёз базального слоя обеспечивает регенерацию эпителия функционального слоя. Усилившийся синтез эстрогенов в яичнике инициирует начало пролиферативной фазы в эндометрии матки. Это приводит к следующим событиям:
эпителиальные клетки базальных отделов маточных желёз делятся митозом, восстанавливают железы и мигрируют к оголённой поверхности эндометрия;
стромальные клетки пролиферируют и секретируют коллаген и основное вещество собственной пластинки слизистой;
спиральные артерии удлиняются по мере восстановления эндометрия.
Пролиферативная фаза продолжается до первого дня после овуляции. К концу этой фазы эндометрий достигает толщины примерно 3 мм, железы узкие, относительно прямые.
В секреторную фазу под воздействием прогестерона происходят драматичные изменения функционального слоя, начинается эта фаза через день или два после овуляции. Эндометрий становится отёчным и постепенно возрастает до 5-6 мм в толщину. Железы увеличиваются, принимают спиралевидную форму, просвет заполняется продуктами их секреции. Слизистая жидкость богата питательными веществами, в особенности гликогеном, необходимым для развития концептуса в случае успешной имплантации. Митозы редки. Рост, наблюдаемый в эту фазы, является результатом гипертрофии эпителиальных клеток, увеличения васкуляризации и отёка эндометрия. Спиральные артерии удлиняются, становятся более закрученными и достигают поверхности эндометрия.
При несостоявшемся оплодотворении жёлтое тело менструации активно продуцирует гормоны в течение 14 дней. Наступающее вслед за этим резкое падение уровня гормонов изменяет кровоснабжение функционального слоя эндометрия, наступает менструальная фаза. Периодические сокращения спиральных артерий приводят к ишемизации функционального слоя, железы прекращают секрецию, эндометрий сокращается в высоту, и строма становится менее отёчной. Через два дня ишемизированный поверхностный эпителий отторгается и прилежащие кровеносные сосуды разрываются. Десквамация эпителия продолжается до тех пор, пока не останется лишь базальный слой.
Так как прямые артерии, питающие базальный слой, не реагируют на изменения гормонального фона, базальный слой остается интактным. Обычно эта фаза продолжается пять дней. Затем цикл повторяется снова.
Шейка матки
Эндометрий шейки матки отличается от эндометрия остальных отделов матки. Слизистая оболочка представлена однослойным цилиндрическим эпителием, его толщина составляет 2-3 мм, и содержит большие разветвлённые цервикальные железы. Спиральные артерии отсутствуют. Слизистая оболочка незначительно изменяется в толщину в разные фазы менструального цикла и не отторгается при менструации, но циклические изменения затрагивают количество и вязкость цервикального секрета, что влияет на транспорт сперматозоидов в цервикальном канале. В середину менструального цикла секреция слизи увеличивается десятикратно, эта жидкая слизь создаёт благоприятные условия для миграции сперматозоидов. В другие фазы цикла вязкая, густая цервикальная слизь препятствует проникновению сперматозоидов в матку.
Нижняя часть шейки матки вдаётся во влагалище (влагалищная часть). Её стенка состоит преимущественно из плотной соединительной ткани с небольшим количеством гладких мышц. Переход однослойного цилиндрического эпителия в многослойный плоский неороговевающий эпителий происходит в отверстии цервикального канала во влагалище – наиболее частое место развития опухолей шейки матки.
ИМПЛАНТАЦИЯ
Имплантация – это процесс проникновения бластоцисты в эндометрий.
Зигота по мере продвижения по маточной трубе проходит серию делений (дробление) без увеличения размеров клеток, что приводит к быстрому увеличению числа клеток. В результате делений образуется морула, каждая клетка которой теперь называется бластомером. На третий день после оплодотворения морула, состоящая из 12-16 клеток и ещё окружённая блестящей оболочкой, достигает полости матки. В течение суток морула плавает в матке, маточная жидкость проникает через блестящую оболочку, образуется заполненная жидкостью полость, бластоцель. Эти события знаменуют образование бластоцисты. Скопление клеток внутри новообразованной полости называется внутренней клеточной массой (эмбриобластом), эти клетки дают начало всем клеткам зародыша; из наружного слоя клеток, трофобласта, в дальнейшем образуется плодовая часть плаценты. В матке бластоциста свободно плавает 1-2 дня до начала имплантации. К этому времени блестящая оболочка растворяется, чтобы клетки трофобласта смогли напрямую контактировать с эндометрием.
При контакте трофобласта (в месте прилегания к нему эмбриобласта) с эндометрием клетки трофобласта прикрепляются к последнему, быстро делятся и дифференцируются с образованием двух слоев:
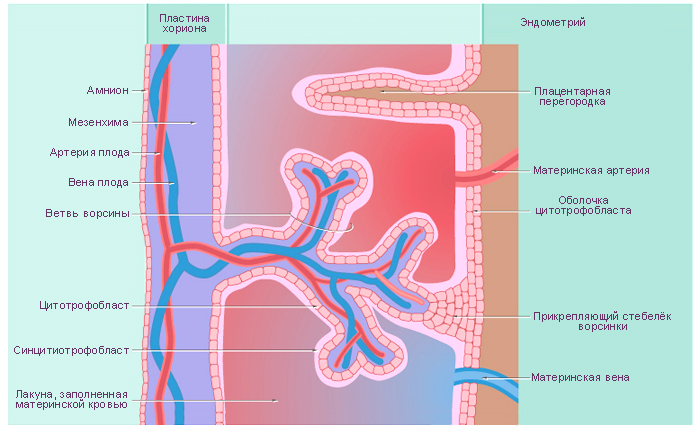
цитотрофобласт – это митотически активный внутренний слой клеток, дочерние клетки которого сливаются вместе с образованием наружного слоя – синцитиотрофобласта;
синцитиотрофобласт состоит из многоядерной массы цитоплазмы; он активно внедряется в эпителий и подлежащую строму эндометрия.
Благодаря ферментной активности трофобласта, бластоциста полностью погружается в эндометрий на 9-й день после оплодотворения. Дальнейшее развитие трофобласта описано в разделе Плацента.
Синцитиотрофобласт имеет хорошо развитый комплекс Гольджи, гладкую и шероховатую эндоплазматическую сеть, многочисленные митохондрии и относительно много липидных капель. Эти морфологические признаки согласуются с функцией этого слоя: секрецией прогестерона, эстрогенов, ХГЧ, человеческого плацентарного лактогена и др.
Изменения эндометрия при имплантации были названы децидуальной реакцией, а эндометрий беременной матки – децидуальной оболочкой. При имплантации бластоцисты под влиянием эстрогенов и прогестерона стромальные клетки эндометрия трансформируются в децидуальные клетки. Образуются крупные, бледные клетки, богатые гликогеном. Хотя точная роль этих клеток не изучена, известно, что они создают благоприятные условия для питания зародыша и предотвращают инвазию трофобласта глубже эндометрия.
Различают три части децидуальной оболочки:
Базальная децидуальная оболочка (decidua basalis) расположена в месте имплантации, она формирует материнскую часть плаценты.
Капсулярная децидуальная оболочка (decidua capsularis) покрывает имплантированную бластоцисту и отделяет её от полости матки.
Париетальная децидуальная оболочка (decidua parietalis) представляет собой остатки эндометрия, т.е. часть, которая напрямую не контактирует с эмбрионом.
К концу третьего месяца внутриутробного развития плод достигает таких размеров, когда капсулярная и париетальная децидуальные оболочки противоположных сторон сливаются, облитерируя полость матки.
Внутренняя клеточная масса формирует двухслойный (биламинарный) зародышевый диск (бластодиск), который собственно и становится эмбрионом, и оболочку внезародышевую мезодерму, которая выстилает внутреннюю поверхность цитотрофобласта. Бластодиск отделяется от внезародышевой мезодермы полостью – внезародышевым целомом. Эмбрион отделяется от эндометрия трёхслойной оболочкой – хорионом. Хорион состоит из производных трофобласта (синцитиотрофобласт и цитотрофобласт) и внутренней клеточной массы (внезародышевая мезодерма).
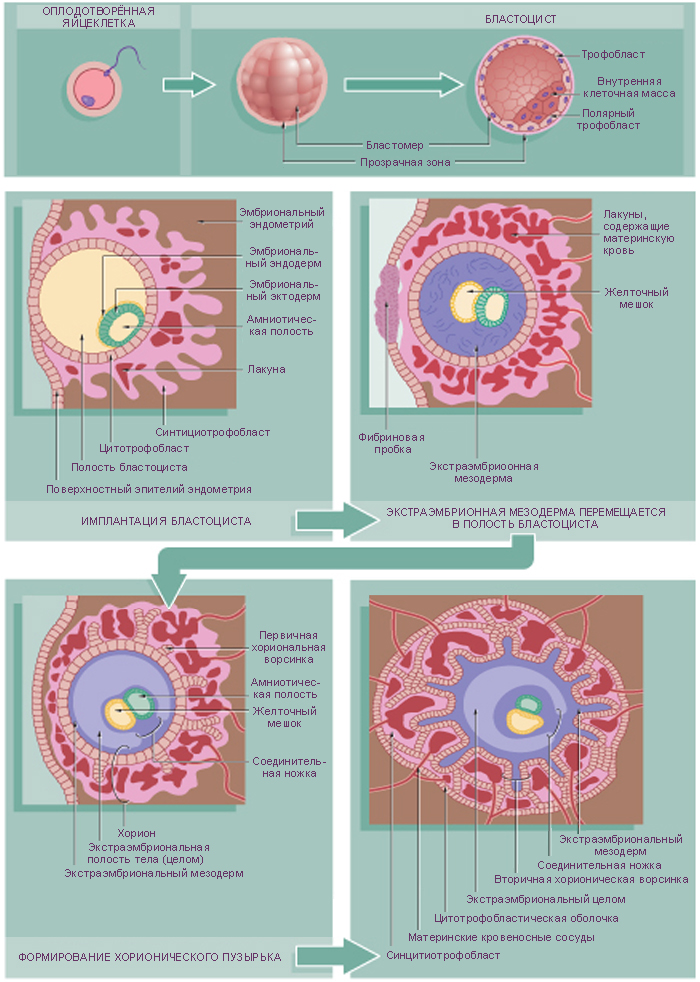
ПЛАЦЕНТА
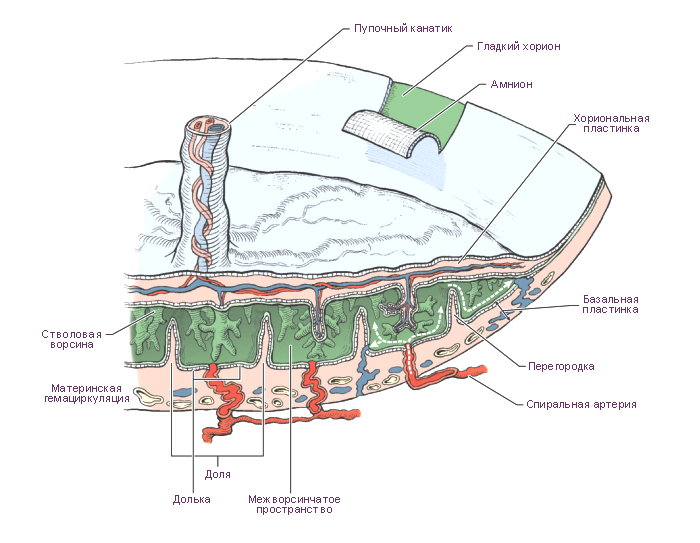
Плацента — это временный орган, который начинает формироваться во время имплантации бластоцисты, и транспортирует питательные вещества и кислород от матери к эмбриону, очищает кровь плода от токсинов и углекислого газа, секретирует гормоны. Плацента состоит из плодового (ворсинчатый хорион) и материнского (базальная децидуальная оболочка) компонентов.
Стадии развития плаценты (плацентация). Погружающийся в стенку матки синцитиотрофобласт окружается маленькими островками эндометрия, содержащими кровеносные сосуды. Ферменты синцитиотрофобласта лизируют материнский эндометрий, оставляя пространства, лакуны, и разрывая кровеносные сосуды. Кровь из повреждённых сосудов заполняет лакуны материнской кровью. Тяжи хорионической ткани (ворсины хориона) прорастают в эти лакуны и развиваются для того, чтобы, во-первых, кровь в сосудах плода и материнская кровь в лакунах находились достаточно близко для полноценного обмена, во-вторых, для формирования избирательно проницаемого плацентарного барьера.
Пролиферация цитотрофобласта, рост внезародышевой мезодермы и кровеносных сосудов последовательно приводят к образованию:
первичных ворсин хориона – скопления клеток цитотрофобласта, окружённого синцитиотрофобластом. Клетки цитотрофобласта имеют неправильную форму и светлую цитоплазму;
вторичных ворсин хориона, которые формируются при прорастании подлежащей внезародышевой мезодермы в первичные ворсины на 12-13 день внутриутробного развития, соответственно они состоят (снаружи внутрь) из синцитиотрофобласта, цитотрофобласта и стержня из внезародышевой мезодермы;
третичных ворсин хориона, в которые, помимо синцитиотрофобласта, цитотрофобласта и внезародышевой мезодермы, врастают кровеносные сосуды (с 3-й недели развития). Клетки внезародышевой мезодермы дифференцируется в эндотелий и образуют кровеносные сосуды, которые позже соединятся с пупочными сосудами плода. К 4-му месяцу цитотрофобласт исчезает, так как все его клетки сливаются с синцитиотрофобластом.
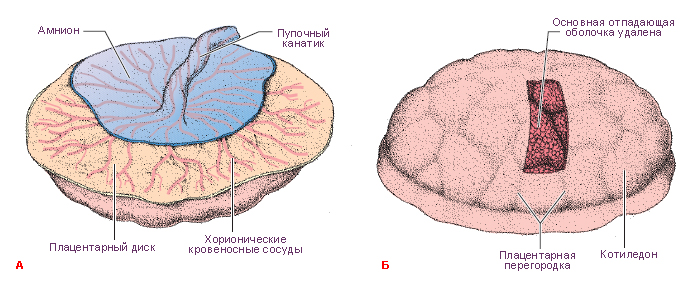
В первые восемь недель развития ворсинки покрывают всю поверхность хориона, но при продолжающемся росте ворсинки на поверхности капсулярной децидуальной оболочки дегенерируют, образуют гладкую, относительно аваскулярную поверхность, называемую гладкий хорион (chorion laeve). Ворсинки, прилежащие к базальной децидуальной оболочке, быстро увеличиваются в размерах, количестве, они разветвляются и составляют ворсинчатый хорион (chorion frondosum), который образует плодовую часть плаценты.
В период бурного роста ворсинчатого хориона на 4-5 месяце гестации плодовая часть плаценты разделяется плацентарными (децидуальными) септами на 15-25 долек, называемых котиледонами. Котиледон – это структурно-функциональная единица сформированной плаценты. Он образован стволовой ворсинкой и её разветвлениями, содержащими сосуды плода. Между котиледонами свободно циркулирует материнская кровь.
Плодовая и материнская кровь не смешивается и отделяется плацентарным барьером. В образовании плацентарного барьера принимают участие ткани плода:
синцитиотрофобласт;
цитотрофобласт (который исчезает к 4-му месяцу гестации);
базальную пластинку трофобласта;
внезародышевую мезенхиму (или мезодерму);
базальную пластинку в стенке капилляра плода;
эндотелий капилляров третичных ворсин хориона.
Функции плаценты.
Транспорт питательных веществ, газов и продуктов метаболизма. К 23-му дню гестации кровь эмбриона циркулирует в третичных ворсинах хориона. Питательные вещества, электролиты, глюкоза, кислород, антитела из материнской крови достигают эмбриона, а позднее – плода, последовательно проходя через плацентарный барьер. Продукты метаболизма плода экскретируются в материнский кровоток.
Синтез гормонов. Плацента – это эндокринный орган. Синцитиотрофобласт хориона синтезирует простагландины, играющие важную роль при начале родовой деятельности, стероидные и пептидные гормоны. Стероидные гормоны прогестерон и эстроген необходимы для поддержания беременности. К плацентарным пептидным гормонам, синтезируемым синцитиотрофобластом и децидуальными клетками, относятся: хорионический гонадотропин человека (ХГЧ), хорионический соматотропин человека (человеческий плацентарный лактоген), инсулиноподобные факторы роста, фактор роста эндотелия, релаксин, лептин, пролактин и др.
ВЛАГАЛИЩЕ
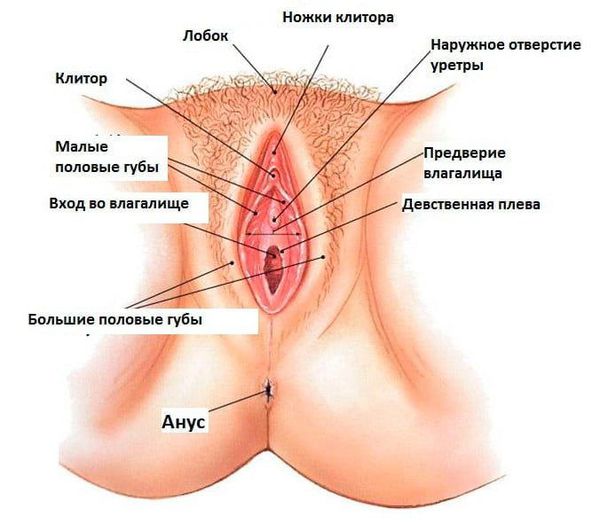
Влагалище представляет собой мышечную трубку, которая соединяет внутренние половые органы с внешней средой. Оно простирается от шейки матки до наружных половых органов. У девственниц влагалище отделяется от наружных половых органов девственной плевой.
Стенка влагалища состоит из трёх слоев:
Слизистая оболочка образует многочисленные поперечные складки и представлена многослойным плоским неороговевающим эпителием и собственной пластинкой слизистой с многочисленными эластическими волокнами. Под действием эстрогенов эпителий влагалища утолщается, эпителиальные клетки накапливают гликоген, который выделяется в просвет. Бактерии в просвете влагалища метаболизируют гликоген, что приводит к накоплению молочной кислоты и низкой pH в просвете органа. Обширное венозное и капиллярное сплетение в собственной пластинке обеспечивает выработку большей части жидкости, которая просачивается в просвет влагалища во время сексуального возбуждения. В вагинальной слизистой содержится немного чувствительных нервных окончаний.
Мышечная оболочка представлена преимущественно продольно ориентированными пучками гладкомышечных клеток (наружный слой), а также мышечными пучками, имеющими циркулярное направление (ближе к слизистой оболочке). Иногда слои плохо различимы друг от друга.
Адвентициальная оболочка. Влагалище окружено футляром из плотной соединительной ткани, содержащем большое количество эластических волокон, обширное венозное сплетение, пучки нервных волокон и скопления нейронов.
В стенке влагалища железы отсутствуют, поверхность многослойного плоского неороговевающего эпителия смачивается секретами цервикальных желёз и больших и малых желёз преддверия. Вагинальная жидкость увеличивает подвижность сперматозоидов.
НАРУЖНЫЕ ПОЛОВЫЕ ОРГАНЫ
Наружные половые органы обозначаются термином вульва и покрыты многослойным плоским эпителием. К вульве относятся:
Клитор является гомологом мужского полового члена и состоит из двух эректильных пещеристых тел, заканчивающихся головкой клитора. Клитор окружен крайней плотью и содержит большое количество чувствительных нервных окончаний.
Преддверие влагалища. В преддверие открываются отверстия уретры и влагалища. Оно содержит 2 типа желез. Большие железы преддверия (Бартолиновы железы) – крупные (размером 1 см), парные трубчато-альвеолярные слизистые железы, расположенные с каждой стороны преддверия, являются аналогами бульбоуретральных желез мужчин. Малые железы преддверия (железы Скина) многочисленные, мелкие слизистые железы, большая часть которых лежит около клитора и наружного отверстия уретры. Эти железы являются аналогами желез Литтре у мужчин.
Малые половые губы. Эти парные складки кожи построены из губчатой (эректильной) соединительной ткани, гомологичной губчатому телу мужского полового члена. Они покрыты многослойным плоским эпителием с малым ороговением, в жировой прослойке стромы расположены многочисленные кровеносные сосуды, эластические волокна и сальные железы, но волосяной покров на них отсутствует.
Большие половые губы представляют собой две продольные кожные складки, гомологичные мошонке у мужчин. Содержат подкожный жир, тонкий слой мышц, сальные и потовые железы. Внутренняя поверхность каждой большой половой губы схожа по строению с малыми половыми губами; на внешней поверхности ороговение выражено больше и имеются грубые волосы.
Наружные половые органы содержат многочисленные чувствительные нервные окончания: тельца Майсснера, тельца Пачини, и свободные нервные окончания.
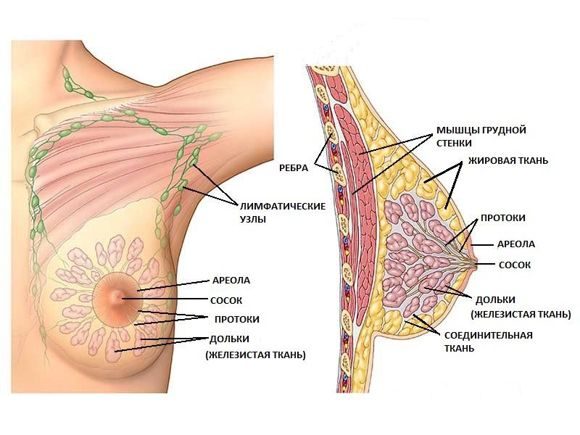
МОЛОЧНЫЕ ЖЕЛЕЗЫ
Молочные железы – это модифицированные апокриновые потовые железы кожи, специализированные секретировать молоко, структура которых значительно изменяется с возрастом, зависит от фазы менструального цикла и репродуктивного статуса женщины. Молочные железы взрослый женщины в неактивном состоянии являются сложными трубчато-альвеолярными железами, состоящими из 15-25 долей, отделённых друг от друга прослойками плотной соединительной ткани. Доли радиально расходятся от соска и делятся на многочисленные дольки. Дольки, основные единицы железы, образуются в пубертатный период под действием прогестерона. Они отделены друг от друга рыхлой соединительной тканью и содержат нескольких секреторных альвеол. Секреторные альвеолы покрыты однослойным кубическим эпителием. В дольках начинаются внутридольковые протоки. Все внутридольковые протоки из одной дольки открываются в один терминальный междольковый проток, который впадает в более крупный млечный проток. Млечный проток каждой доли открывается отдельным отверстием на вершине соска. Под ареолой, пигментированной областью вокруг соска, каждый млечный проток имеет терминальное расширение – млечный синус.
Эпителиальная выстилка протоков постепенно меняется с многослойного плоского ороговевающего эпителия (у отверстий млечных протоков) на двухслойный кубический (млечный синус) и, в конечном итоге, на однослойный цилиндрический или кубический эпителий (на остальном протяжении протоков) с плотно прилегающими миоэпителиальными клетками, отделяющими эпителий от базальной мембраны. Сосок и ареола выстланы многослойным плоским ороговевающим эпителием.
В ареоле сосков располагаются сальные, потовые железы и модифицированные молочные железы (железы Монтгомери). Эти железы структурно занимают промежуточное положение между потовыми и истинно молочными железами. Они незначительно выступают над поверхностью ареолы, и продуцируют секрет, увлажняющий эпидермис, изменяющий рН кожи и предотвращающий рост микробов.
Эмбриональное развитие. В эмбриогенезе молочные железы закладываются у зародышей обоих полов. Зачатки молочных желёз развиваются вдоль молочных линий (валиков) – парных утолщений эктодермы, появляющихся на 6-й неделе развития на переднебоковых стенках тела между развивающимися подмышечной и паховой областями. Очень быстро почти на всём протяжении молочные линии подвергаются регрессу, кроме одной пары на груди.
Позднее 15-25 эпителиальных инвагинаций пенетрируют в подлежащую мезенхиму. Это будущие млечные протоки. Секреция молочных желез (под влиянием материнских гормонов и гормонов плаценты) встречается у новорожденных.
До начала пубертатного периода женские и мужские молочные железы у детей обоих полов развиваются схожим образом. Ювенильная молочная железа неактивна, состоит из млечных протоков и синусов, погружённых в соединительную ткань. Маленький сосок окружен бледно-пигментированным околососковым кружком – ареолой. У мальчиков на заре полового созревания тестостерон воздействует на мезенхимальные клетки молочных желёз, подавляя их рост. В то же время у девочек молочных железы подвергаются дальнейшему развитию под действием эстрогена и прогестерона.
Изменения во время полового созревания. Половое созревание и синергичное влияние эстрогенов, гормона роста, пролактина и глюкокортикоидов стимулирует дальнейшее развитие выводных протоков и накопление жировой ткани. Женская грудь увеличивается в размерах, соски становятся более выступающими. Усиленный синтез эстрогенов яичниками стимулирует рост млечных протоков (увеличение их длины, диаметра и ветвления).
Строение нелактирующей молочной железы описано выше. Незначительные изменения происходят во время менструального цикла. Пик эстрогенов во время овуляции индуцирует дальнейшую пролиферацию клеток протоков, что может вызвать предменструальное увеличение желез с сопутствующим преходящим отёком и повышенной чувствительностью.
Молочные железы во время беременности. Влияние нескольких гормонов, включая эстрогены, прогестерон, пролактин и плацентарный лактоген человека, вызывает интенсивную пролиферацию протоков и рост концевых участков альвеол, приводя к увеличению железы. Эпителий внутридольковых протоков пролиферирует, дифференцируется в клетки, продуцирующие молоко, и образует многочисленные секреторные альвеолы внутри долек. Во время беременности ядро альвеолярной клетки смещается к базальному полюсу, сверху его окружают комплекс Гольджи, митохондрии, лизосомы, секреторные везикулы с молочным белком и несколько апикально расположенных капель жира. Миоэпителиальные клетки лежат между альвеолярными клетками и их базальной мембраной. На поздних сроках беременности возрастает число плазматических клеток в междольковой соединительной ткани. Эти клетки добавляют секреторный IgA к секрету молочных желез (особенно к молозиву) и обеспечивают пассивный иммунитет новорожденного. Несмотря на то, что железы хорошо развиваются при беременности, секреции в них не наблюдается вплоть до поздних сроков беременности, когда они содержат молозиво (первое молоко с высоким содержанием белка), или до лактации, когда они содержат богатое жирами молоко.
Лактирующая молочная железа. После рождения плаценты уровни эстрогенов и прогестерона падают, а пролактина возрастают. Главное гистологическое отличие лактирующей молочной железы от железы во время беременности заключается в наличии молока в просвете альвеол и их сопутствующая дилятация. Секреторные клетки уменьшаются по высоте от цилиндрического до кубического эпителия, в их цитоплазме обнаруживаются многочисленные капельки жира, содержащие нейтральные триглицериды. Капельки жира секретируются вместе с фрагментами клеточной мембраны и цитоплазмы (апокриновая секреция). Секреторные пузырьки с молочными белками освобождаются путём экзоцитоза (мерокриновая секреция).
Состав молока. Молоко содержит 4 % липидов, 1,5 % белков
(казеины, лактоальбумин, IgA), 7 % лактозы (дисахарид глюкозы и галактозы) и 87,5 % воды.
Поддержание лактации. При сосании чувствительные стимулы ингибируют секрецию допамина (пролактин-ингибирующего гормона) гипоталамусом. Это приводит к выбросу пролактина из передней доли гипофиза, что, в свою очередь, стимулирует продукцию молока. При отнятии ребенка от груди и прекращении сосания уровень пролактина падает и альвеолы дегенерируют.
Рефлекс выделения молока. Чувствительный стимул при сосании вызывает синтез окситоцина в гипоталамусе. При выделении окситоцина задней долей гипофиза миоэпителиальные клетки сокращаются, изгоняя молоко из альвеол в млечные протоки.
Старческая инволюция. После менопаузы на фоне снижения секреции яичниками половых гормонов секреторные отделы, протоки, жировая и междольковая соединительная ткани молочной железы атрофируются.
Пингбэк: Половая система — yavmede.ru