ГАМЕТОГЕНЕЗ
Мужские половые клетки (сперматозоиды)
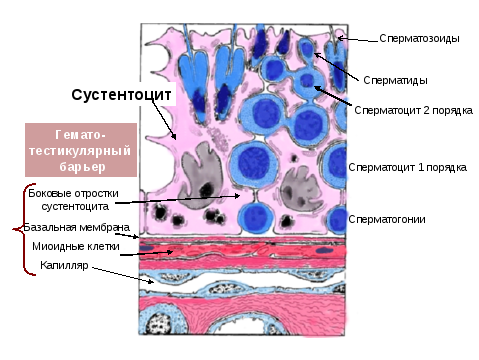
Сперматогенез (процесс образования сперматозоидов из сперматогоний) происходит в извитых семенных канальцах яичка, многослойный эпителий которых состоит из сперматогенных клеток и поддерживающих клеток Сертоли.
Первичные половые клетки (гоноциты) мигрируют из энтодермы желточного мешка в гонадный валик на задней стенке брюшной полости и начинают сложный многоступенчатый процесс дифференцировки, который заканчивается выходом сперматозоидов в просвет канальца.
Сперматогонии. Это мелкие округлые клетки, лежащие рядом с базальной пластинкой извитого семенного канальца. До начала полового созревания это единственный тип сперматогенных клеток, присутствующих в извитых канальцах. Это наименее дифференцированные клетки с круглым ядром и неоднородным гетерохроматином. Клетки имеют диплоидный набор хромосом (46, 2n) и диплоидное количество ДНК (2N).
Первичные сперматоциты. Эти клетки находятся ближе к просвету канальца, чем сперматогонии. Это самые крупные клетки, они имеют большое круглое ядро с темными пучками гетерохроматина. В них обычно происходит профаза I деления мейоза. Эта фаза мейоза самая длинная, она продолжается 22 дня. Клетки имеют диплоидный набор хромосом (46, 2n) и тетраплоидное количество ДНК (4N).
Вторичные сперматоциты находятся еще ближе к просвету канальца. Образуются в результате первого мейотического деления первичных сперматоцитов, и в 2 раза меньше последних по размеру. На гистологических препаратах видны редко, так как вступают во второе мейотическое деление почти сразу после образования. Клетки имеют гаплоидный набор хромосом (23, n) и диплоидное количество ДНК (2N).
Сперматиды. Это продукт второго мейотического деления вторичных сперматоцитов, расположены близко к просвету канальцев. Это мелкие клетки с тёмным гетерохроматиновым ядром. Ядро может иметь разную морфологию в зависимости от стадии, на которой находится клетка. Клетки имеют гаплоидный набор хромосом (23, n) и гаплоидное количество ДНК (N).
Сперматозоиды расположены в просвете канальца. Это результат сперматогенеза, продукт дифференцировки сперматид. Отличительный признак – длинный жгутик. Клетки имеют гаплоидный набор хромосом (23, n) и гаплоидное количество ДНК (N).
Сперматогенез
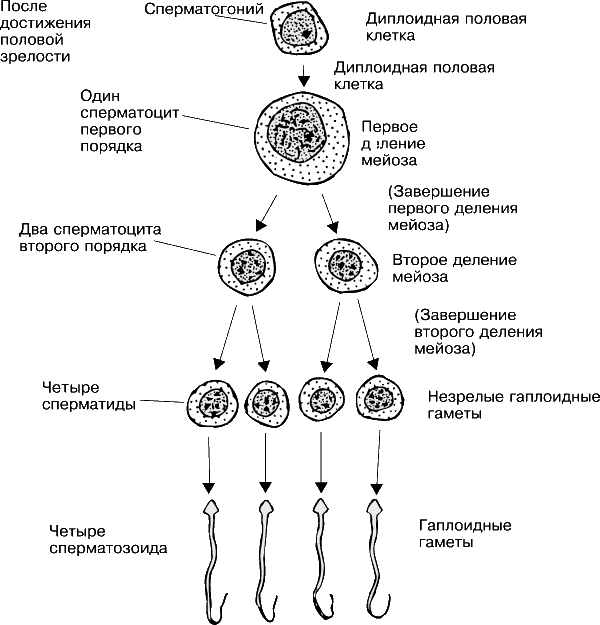
Сперматогенез – процесс, который происходит под действием тестостерона и состоит из трёх этапов: сперматоцитогенеза, мейоза и спермиогенеза.
1. Сперматоцитогенез. Это процесс образования первичных сперматоцитов из сперматогониев путём серии стандартных митотических делений. При этом в результате ранних делений образуются сперматогонии двух типов, А и В:
сперматогонии А остаются недифференцированными клетками, способными и далее продуцировать сперматогонии А и В;
сперматогонии В могут подвергаться митозу и продуцировать вновь сперматогонии В или первичные сперматоциты, вступающие в мейоз.
2. Мейоз. Это процесс двух последовательных клеточных делений, в результате которых образуются 4 гаплоидных клетки (сперматиды) из диплоидных первичных сперматоцитов.
Мейоз I. В этом делении из первичных сперматоцитов образуются вторичные сперматоциты.
S-фаза (синтез ДНК) – перед делением ДНК удваивается, и клетка становится тетраплоидной.
В профазе 23 пары удвоенных гомологичных хромосом утолщаются и скручиваются, соединяются в определённых точках (формируют мостики (хиазмы)) и ДНК перемещается между гомологичными хромосомами
(кроссинговер). В конце профазы исчезает ядерная мембрана.
В метафазе 23 пары удвоенных хромосом (биваленты) выстраиваются на экваторе клетки.
В анафазе гомологичные хромосомы (каждая состоит из 2-х хроматид) каждой пары расходятся к противоположным полюсам клетки. Центромеры не расщепляются, поэтому сестринские хроматиды продолжают удерживаться вместе. Вследствие кроссинговера сестринские хроматиды уже могут быть неидентичными. Таким образом, к полюсам движутся хромосомы, состоящие из двух хроматид. В ходе этой фазы из одной диплоидной клетки образуется две гаплоидные (но с диплоидным количеством ДНК).
В телофазе у дочерних клеток образуется ядерная мембрана. Каждая клетка – гаплоидная, содержит одну из удвоенных гомологичных хромосом (каждая состоит из 2-х хроматид, т.е. количество ДНК – диплоидное).
Мейоз II. В этом делении из вторичных сперматоцитов образуются сперматиды. В анафазе этого деления хроматиды каждой хромосомы расходятся к противоположным полюсам клетки. Таким образом, число хромосом в каждой новой клетке остается гаплоидным, количество ДНК уменьшается вдвое (гаплоидное).
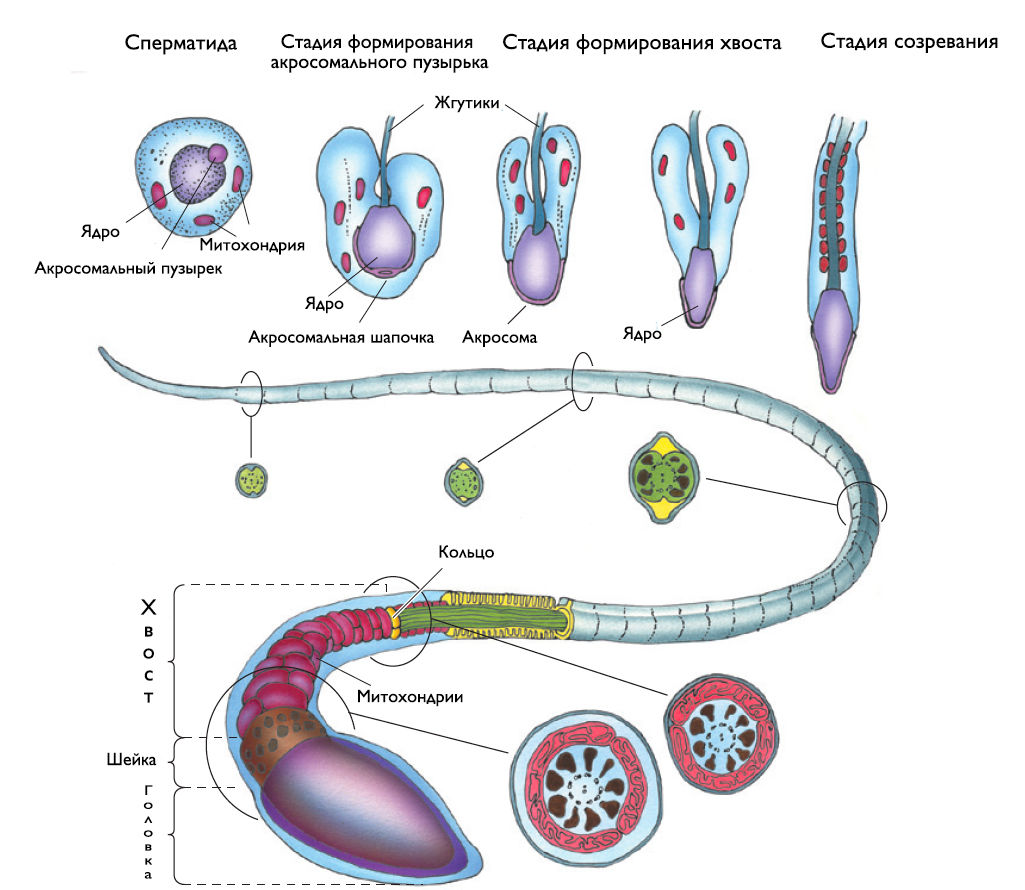
3. Спермиогенез. Это сложный процесс клеточной дифференцировки, в результате которого сперматида становится сперматозоидом (рис. 2). Этапы:
Формирование акросомы. Проакросомальные гранулы образуются в комплексе Гольджи, сливаются и формируют большую акросомальную везикулу, которая двигается к ядру и прикрепляется к его оболочке. Акросома покрывает как шапочка почти половину ядра. Она содержит гидролитические ферменты (гиалуронидаза, нейраминидаза, кислая фосфатаза, трипсин и т.д.), которые растворяют оболочку яйцеклетки при оплодотворении.
Формирование жгутика. Центриоли мигрируют к заднему полюсу сперматиды, и от одной из них начинает «расти» хвост сперматозоида, а другая формирует футляр вокруг его основания.
Миграция митохондрий. Митохондрии двигаются к жгутику и формируют спиральный футляр вокруг проксимальной части хвоста (в средней части), концентрируясь в месте будущего большого потребления энергии. Фруктоза и лимонная кислота (митохондриальные метаболиты) спермы необходимы для движения сперматозоидов.
Конденсация хроматина. Хроматин формирует плотную массу с невидимой структурой. Цилиндрический пучок микротрубочек (манжета) окружает ядро, соединяясь с задним краем акросомы, что приводит к уплощению и удлинению ядра.
Удаление цитоплазмы. При выходе сперматозоида в просвет канальца он теряет остатки цитоплазмы, которая фагоцитируется клетками Сертоли.
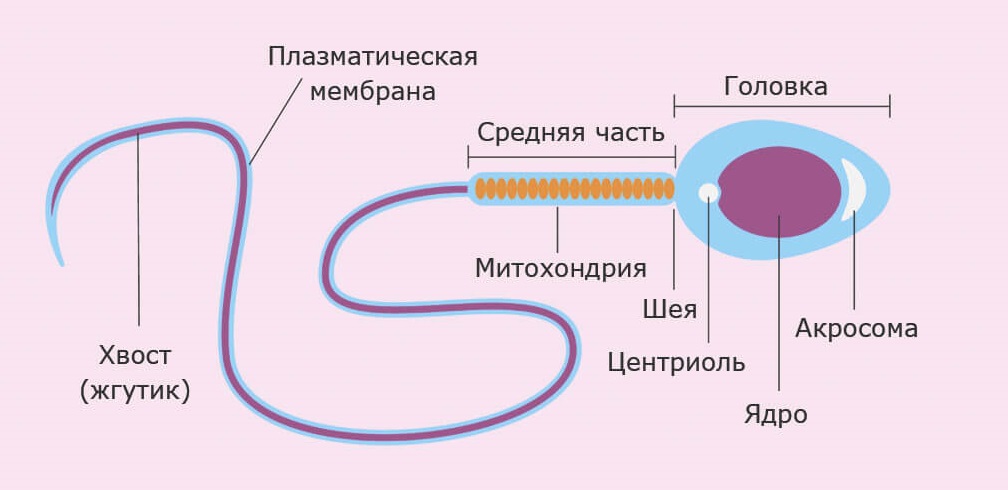
Строение сперматозоида
Сперматозоид человека состоит из двух основных частей: головки и хвоста. Головка содержит конденсированное ядро с гаплоидным набором хромосом, акросому и тонкий слой цитоплазмы. В хвосте выделяют 4 части:
связующий отдел, или шейка, содержит центриоль;
промежуточный отдел содержит большое количество митохондрий;
главный отдел;
конечный (терминальный) отдел.
Женские половые клетки (яйцеклетки)
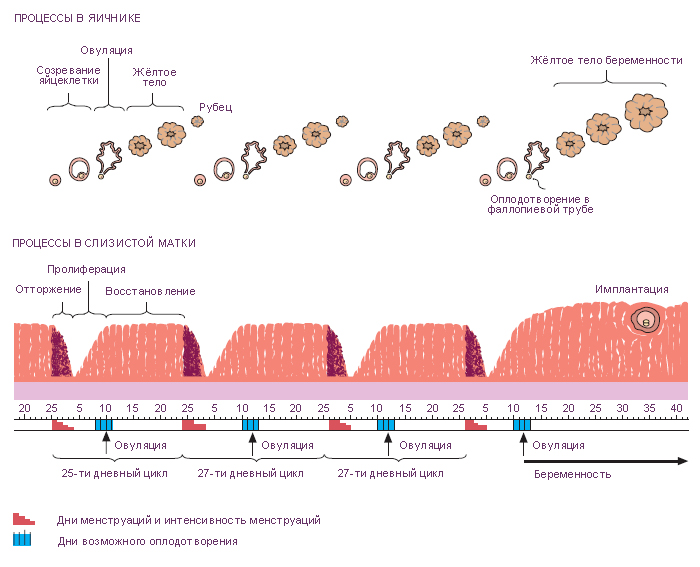
Яйцеклетки (ооциты) находятся в специальных пузырьках – фолликулах яичника.
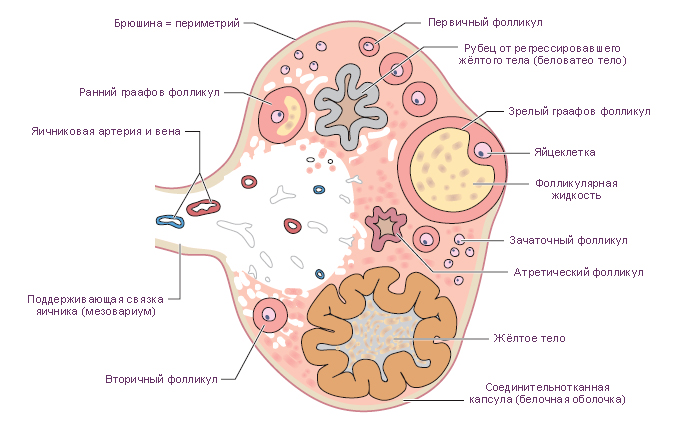
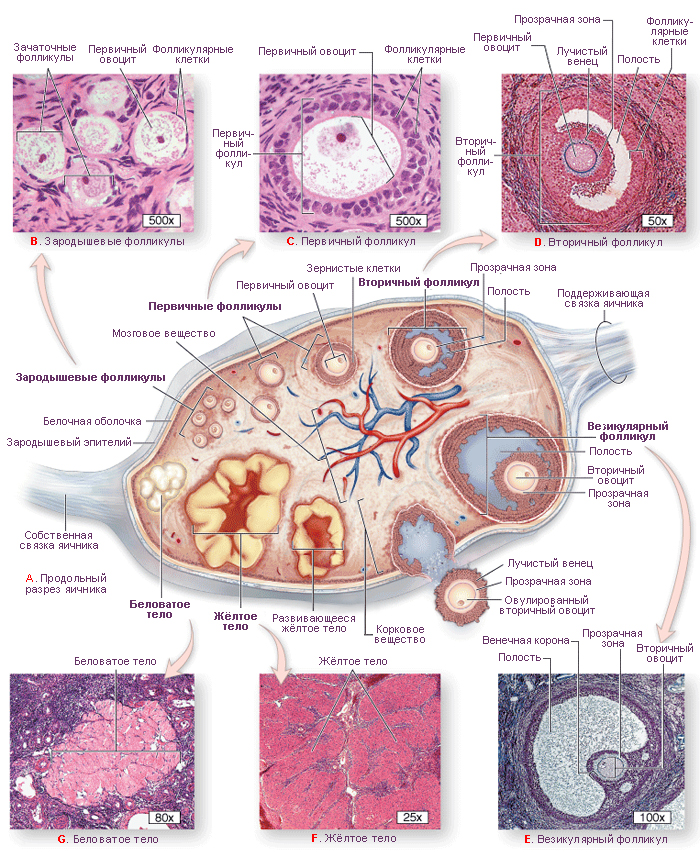
Фолликулы яичника. Каждый фолликул содержит один ооцит, окружённый одним или несколькими слоями фолликулярных (гранулёзных) клеток. В корковом веществе находятся фолликулы на разных стадиях развития.
1. Примордиальные фолликулы. Это самая ранняя стадия развития фолликула. До начала полового созревания в яичнике девочки есть только такие фолликулы. Каждый такой фолликул содержит первичный ооцит (большинство находится в профазе I мейоза), окружённый одним слоем плоских фолликулярных клеток (рис. 4А).
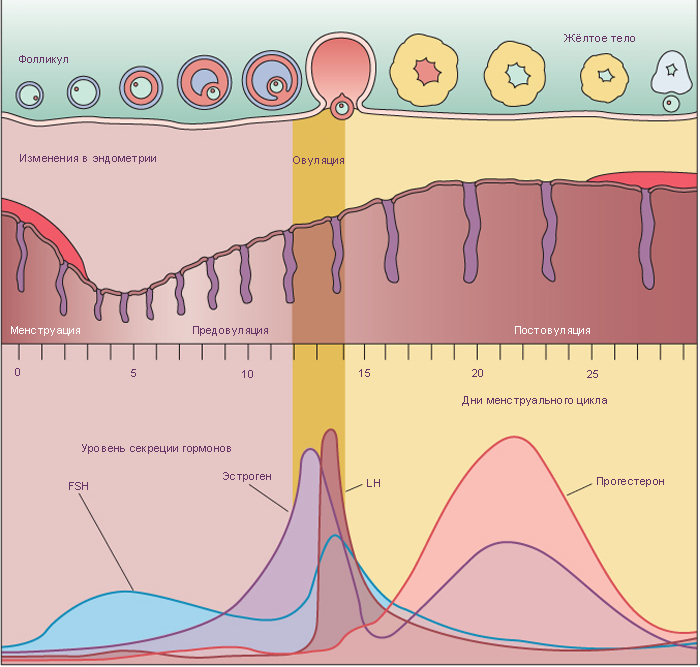
2. Растущие фолликулы
Фолликулы начинают расти под действием фолликул-стимулирующего гормона гипофиза (ФСГ). Ооцит увеличивается в диаметре до 125-150 мкм. Фолликулярные клетки становятся кубическими и пролиферируют, в результате чего фолликулярный эпителий становится многослойным. Соединительная ткань, непосредственно окружающая фолликул, образует теку (оболочку) фолликула, клетки которой продуцируют стероидные гормоны.
Первичные фолликулы – первичный ооцит окружён одним слоем кубических фолликулярных клеток. Полость отсутствует. На этой стадии между ооцитом и фолликулярными клетками образуется блестящая оболочка (zona pellucida), богатая гликопротеинами ZP-1, ZP-2, ZP-3. Самый важный гликопротеин ZP-3 является рецептором, связывающим сперматозоид и запускающим акросомную реакцию; предполагается, что ZP-2 является вторичным рецептором сперматозоидов, а функциональная роль ZP-1 ещё не была охарактеризована.
Вторичные фолликулы. После быстрых митотических делений из одного слоя фолликулярных клеток образуется многослойный кубический эпителий. Базальная мембрана смещается и располагается между наружными слоями фолликулярных клеток и соединительнотканной стромой. Между фолликулярными клетками формируются многочисленные щелевые контакты для обмена питательными веществами и информационными макромолекулами. Во время пролиферации фолликулярных клеток, стромальные клетки, окружающие фолликул, формируют новую оболочку, называемую текой фолликула. Эта оболочка располагается кнаружи от базальной мембраны.
Третичные (антральные) фолликулы характеризуются наличием заполненной жидкостью полости. Из фолликулярных клеток формируется гранулёза (фолликулярные клетки с этого момента называются гранулёзными клетками). Гранулёзные клетки начинают секретировать фолликулярную жидкость, в результате чего между ними образуются заполненные жидкостью мелкие полости, которые постепенно сливаются в одну большую полость – антрум. Первичный ооцит смещается к одному из полюсов фолликула, увеличивается до 125-150 мкм в диаметре и перестаёт расти, фолликул же достигает 10 мм и более в диаметре. Тека фолликула разделяется на 2 слоя: внутренняя тека содержит богатую сосудистую сеть и текальные эндокринные клетки с хорошо развитой гладкой ЭПС, секретирующие андрогены (последние транспортируются к гранулезным клеткам, которые с помощью ароматазы превращают андрогены в эстрогены). Наружная тека представлена, главным образом, богатой сосудами соединительной тканью, содержащей текальные фибробласты.
- Зрелые (Граафовы) фолликулы отличаются от третичных фолликулов, главным образом, своими крупными размерами (2,5 см в диаметре). В эту стадию: 1) полость фолликула (антрум) значительно увеличивается в размерах; 2) ооцит смещается к одной из сторон фолликула; 3) вокруг ооцита образуется лучистый венец (corona radiata) из нескольких слоёв гранулёзных клеток; 4) в месте прилегания ооцита к гранулёзному слою из последних формируется яйценосный бугорок (cumulus oophorus) – «пьедестал» ооцита. Эта стадия предшествует овуляции. Примерно за 24-36 ч до овуляции первичный ооцит завершает первое деление мейоза, что в итоге приводит к образованию вторичного ооцита и первого полярного тельца.
3. Атретические фолликулы. Хотя при рождении в яичниках имеется около 1 млн фолликулов, только 450 достигают зрелости. Более 99 % становятся атретическими, т.е. они дегенерируют путем аутолиза на разных стадиях развития.
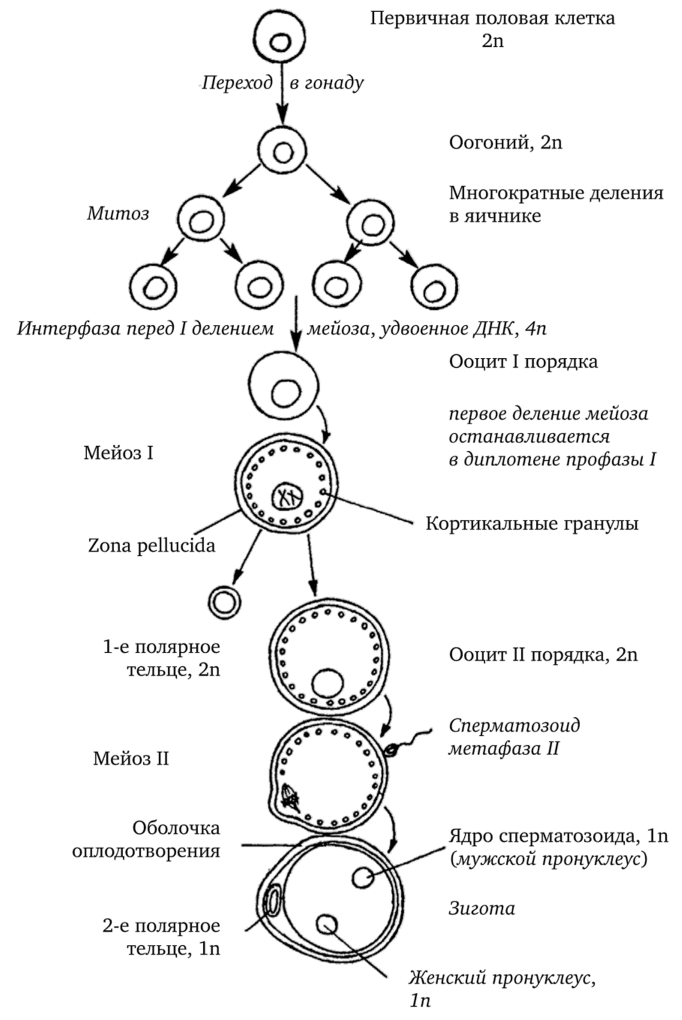
Оогенез
Эндодерма желточного мешка дает начало примордиальным половым клеткам, которые мигрируют в половые валики на задней стенке брюшной полости, из которых развиваются яичники.
Половые клетки окружены уплощёнными фолликулярными клетками примордиальных фолликулов; они вступают в первое деление мейоза и останавливаются в профазе. На этом этапе они называются первичными ооцитами (сравнимы с первичными сперматоцитами).
Первое деление мейоза заканчивается непосредственно перед овуляцией, при этом происходит равномерное деление хроматина и неравное деление цитоплазмы между образовавшимися дочерними клетками. Первая клетка с большим количеством цитоплазмы называется вторичным ооцитом; вторая клетка с меньшим количеством цитоплазмы называется первым полярным тельцем.
После своего формирования, но до начала овуляции, вторичный ооцит вступает во второе мейотическое деление, которое останавливается в метафазе до наступления оплодотворения. В момент оплодотворения второе деление мейоза заканчивается и образуется яйцеклетка и второе полярное тельце. Оплодотворённая яйцеклетка называется зиготой.
Строение яйцеклетки
Яйцеклетка состоит из ядра с гаплоидным набором хромосом и выраженной цитоплазмы, в которой содержатся все органеллы, за исключением клеточного центра.
Выделяют три оболочки, покрывающих яйцеклетку снаружи:
плазматическая мембрана яйцеклетки (оолемма);
блестящая оболочка (zona pellucida)
лучистый венец (corona radiata), образованный гранулёзными клетками фолликула.
Перивителлиновое пространство располагается между блестящей оболочкой и плазматической мембраной ооцита. В этом пространстве находятся первое, а в случае оплодотворения и второе, полярные тельца.
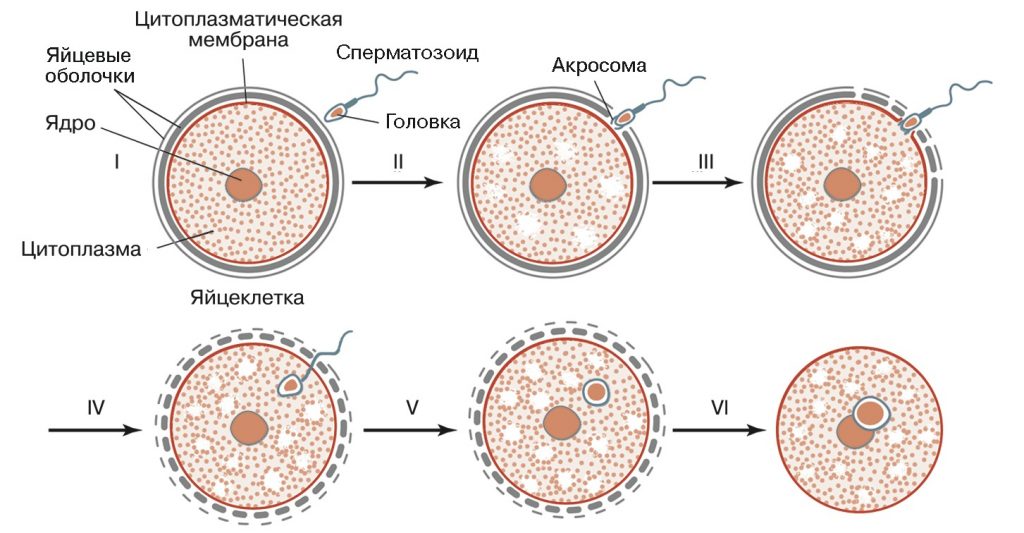
ОПЛОДОТВОРЕНИЕ
Оплодотворение в норме происходит в ампуле маточной трубы.
Только что эякулированные сперматозоиды к оплодотворению вторичного ооцита не способны. Они должны подвергнуться процессу капацитации до того, как приобретут способность прободать оболочки яйцеклетки. Капацитация обычно осуществляется внутри женских репродуктивных путей, хотя этот процесс можно индуцировать in vitro в соответствующих условиях культивирования.
Капацитация – это процесс активации сперматозоидов, приобретения ими оплодотворяющей способности, который включает в себя структурные и функциональные изменения сперматозоидов. В процессе капацитации сначала удаляется гликопротеиновый слой с плазматической мембраны сперматозоида, что в последующем приводит к реорганизации поверхностного слоя клеточной мембраны. У капацитированных сперматозоидов изменяются движения жгутика с регулярных волнообразных на хлыстоподобные. Такое биение жгутика способствует более эффективному движению сперматозоидов. Капацитированные сперматозоиды приобретают повышенную чувствительность к ионам кальция и имеют большее содержание цАМФ, что необходимо для осуществления следующего этапа процесса оплодотворения – акросомной реакции. Капацитация занимает несколько часов.
Акросомная реакция очень важна для оплодотворения, так как без неё сперматозоид не может пройти через блестящую оболочку яйцеклетки. До начала акросомной реакции сперматозоид прободает лучистый венец. Акросомная реакция начинается с контакта сперматозоида с ZP-3 рецепторами блестящей оболочки. В результате ферменты акросомы (в основном гиалуронидазы) растворяют блестящую оболочку. После этого мембрана сперматозоида сливается с мембраной яйцеклетки, и ядро сперматозоида проникает в цитоплазму вторичного ооцита.
Проникновение ядра сперматозоида в цитоплазму вторичного ооцита приводит к завершению второго деления мейоза. В результате образуется яйцеклетка и второе полярное тельце.
После проникновения одного сперматозоида в яйцеклетке возникает кортикальная реакция. Она заключается в следующем. В цитоплазму яйцеклетки (ооплазму) входят ионы натрия, в результате чего меняется заряд оолеммы с отрицательного на положительный. Кроме того, в ооплазме резко повышается концентрация ионов кальция. Всё это приводит к тому, что кортикальные гранулы начинают двигаться к оолемме и происходит экзоцитоз содержимого кортикальных гранул. Ферменты кортикальных гранул разрушают рецепторы для сперматозоидов и изменяют свойство блестящей оболочки, в результате чего другие сперматозоиды уже не могут проникнуть в ооплазму. Оолемма и блестящая оболочка яйцеклетки с видоизмененными свойствами получают название оболочки оплодотворения. После проникновения сперматозоида в яйцеклетку ядра этих клеток сначала располагаются по отдельности (стадия двух пронуклеусов), а потом сливаются, образуя синкарион.
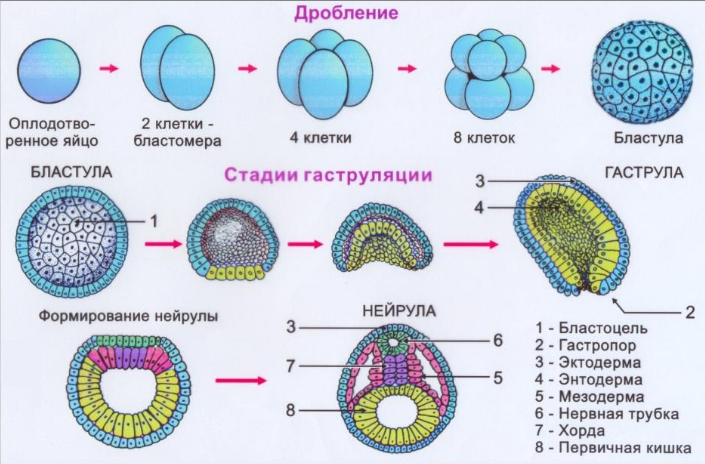
ДРОБЛЕНИЕ
Оплодотворение происходит в ампуле маточной трубы, в результате гаплоидные мужской и женский пронуклеусы сливаются с образованием диплоидного ядра зиготы.
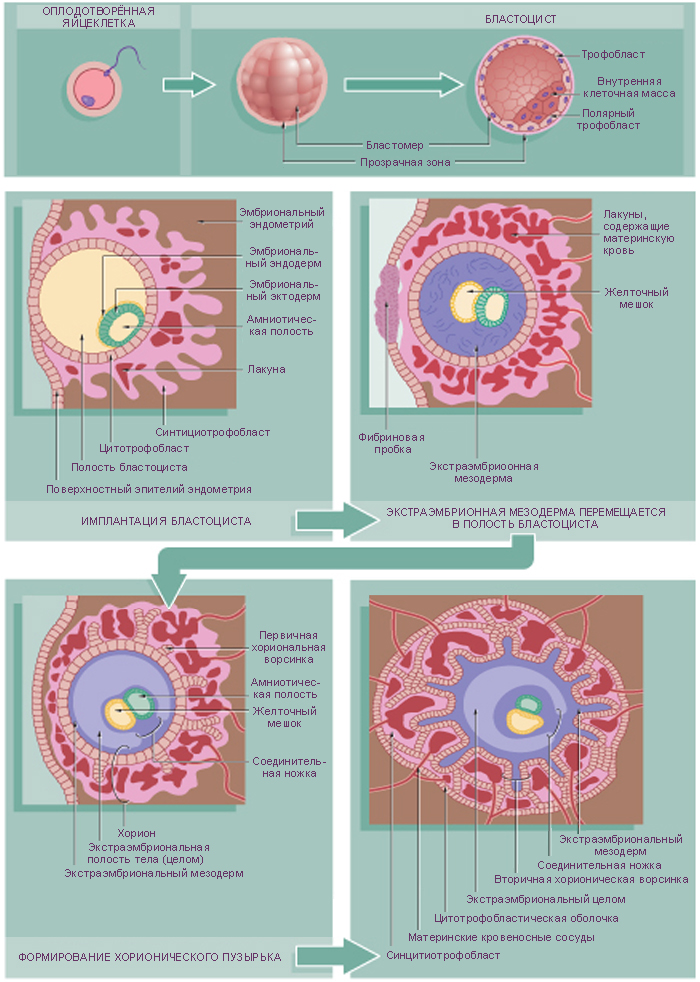
Дробление – митотическое деление диплоидных клеток без увеличения их суммарного объема. При дроблении происходит относительно быстрое увеличение количества клеток, при этом клетки уменьшаются в размерах, таким образом, диаметр концептуса не изменяется. Эти клетки были названы бластомерами. Первое дробление начинается примерно через 30 часов после оплодотворения (Рис. 1). Плоскость первого деления проходит через полярные тельца (которые в этой связи именуются направительными).
Условно на стадии 8-ми бластомеров происходит компактизация концептуса, в результате чего поверхность соприкосновения бластомеров увеличивается, а объем межклеточного пространства уменьшается. Компактизация создает условия, обеспечивающие первый шаг к дифференцировке – обособлению наружной клеточной массы (трофобласта) от внутренней клеточной массы.
Дробление у человека голобластическое (то есть зигота полностью разделяется на два бластомера), асинхронное (разные бластомеры дробятся с различной скоростью, поэтому зародыш на отдельных стадиях дробления содержит нечетное число клеток) и неравномерное (в результате дробления образуются бластомеры неравной величины). При дроблении зигота, а затем и морула, окружена оболочкой оплодотворения, сохраняющейся до стадии бластоцисты.
Дробление отличается от обычного клеточного цикла отсутствием фаз G1 и G2 и очень короткой S фазой, во время которой происходит синтез ДНК. На стадии бластоцисты дробления прекращаются и клетки возобновляют обычный клеточный цикл.
МОРУЛА
По мере увеличения числа бластомеров концептус перемещается по маточной трубе и к третьим суткам достигает стадии морулы.
Морула – это группа клеток (стадия 16 бластомеров), возникших в ходе нескольких делений дробления и заключённых внутри оболочки оплодотворения (Рис. 2, С). Центрально расположенные клетки морулы соединяются щелевыми контактами. Именно из этих клеток и возникает собственно эмбрион. Периферические клетки морулы соединяются плотными контактами. Из них в последующем образуется трофобласт, а затем и плацента.
БЛАСТОЦИСТА
К 4-5-ым суткам после оплодотворения морула попадает в матку. В периферических клетках открываются натриевые каналы, и вслед за ионами натрия в концептус устремляется вода из полости матки. Сначала жидкость накапливается между клеткми и образует небольшие полости, которые затем сливаются в единую полость – бластоцель. На этой стадии развития зародыш носит название бластоцисты. Бластоцисту составляет трофобласт и внутренняя клеточная масса (эмбриобласт).
Трофобласт – наружная оболочка зародыша, которая образована периферическими клетками, из трофобласта в дальнейшем образуется плодовая часть плаценты. Крупные удлиненные клетки трофобласта соединены при помощи плотных контактов, эти клетки «накачивают» жидкость в бластоцель.
Внутренняя клеточная масса (эмбриобласт) – компактная масса мелких клеток, выступающих в бластоцель. Клетки эмбриобласта происходят из центральной части морулы. В дальнейшем из эмбриобласта образуется собственно зародыш и некоторые связанные с ним оболочки. Клетки эмбриобласта тотипотентны, то есть являются эмбриональными стволовыми клетками.
В матке бластоциста свободно плавает 2-3 дня до начала имплантации. К этому времени оболочка оплодотворения истончается и исчезает, чтобы клетки трофобласта смогли напрямую контактировать с эндометрием (внутренним слоем матки). Оболочка оплодотворения предотвращает имплантацию бластоцисты в маточной трубе.
ИМПЛАНТАЦИЯ
Имплантация – это проникновение бластоцисты в эндометрий. Этот этап включает в себя важные изменения бластоцисты и эндометрия (децидуальная реакция, которая будет описана ниже).
Процесс имплантации подразделяется на две фазы: 1) фазу адгезии — прилипания к эндометрию; и 2) фазу инвазии —внедрения в эндометрий.
Процесс имплантации происходит на 7-8-е сутки и продолжается в течение 40 ч.
Клетки трофобласта прикрепляются к эндометрию, быстро делятся и дифференцируются с образованием двух слоев. Синцитиотрофобласт – инвазивный наружный слой, который представляет собой многоядерную клеточную структуру, без четких границ между клетками. Образуется он в результате слияния одноядерных клеток, выталкиваемых из внутреннего слоя – цитотрофобласта. Синцитиотрофобласт разрушает эндометрий ферментами, благодаря чему эмбрион может внедряться в строму матки.
На 9-й день после оплодотворения дефект эпителия в месте внедрения зародыша зарастает, и зародыш оказывается окруженным со всех сторон трофобластом и лакунами (или полостями), заполненными материнской кровью, изливающейся из разрушенных сосудов эндометрия. Во время имплантации зародыша происходят изменения не только в трофобласте, но и в эмбриобласте, где происходит гаструляция.
В процессе имплантации происходит смена гистиотрофного типа питания зародыша на гематотрофный.
С момента оплодотворения и включая имплантацию, питание зародыша осуществляется за счет секрета маточных труб и продуктов распада эндометрия. Такой тип питания – гистиотрофный.
После разрушения стенки сосуда ворсины трофобласта омываются кровью, содержащейся в лакунах, и питание зародыша осуществляется за счет усвоения трофобластом питательных веществ, растворенных в крови матери. Такой тип питания – гематотрофный.
Период имплантации является первым критическим периодом развития зародыша.
ГАСТРУЛЯЦИЯ
После образования бластоцисты начинается гаструляция – процесс образования трёх зародышевых листков (эктодермы, мезодермы и энтодермы).
Для удобства рассмотрения хода и результатов гаструляции ее условно разделяют на две фазы. Первая фаза гаструляции протекает на 7-8-е сутки (в процессе имплантации) и осуществляется способом деламинации (формируется эпибласт, гипобласт). Вторая фаза гаструляции происходит с 14-х на 17-е сутки.
В период между I и II фазами гаструляции, т. е. с 9-х по 14-е сутки формируются внезародышевая мезодерма и три внезародышевых органа — хорион, амнион и желточный мешок.
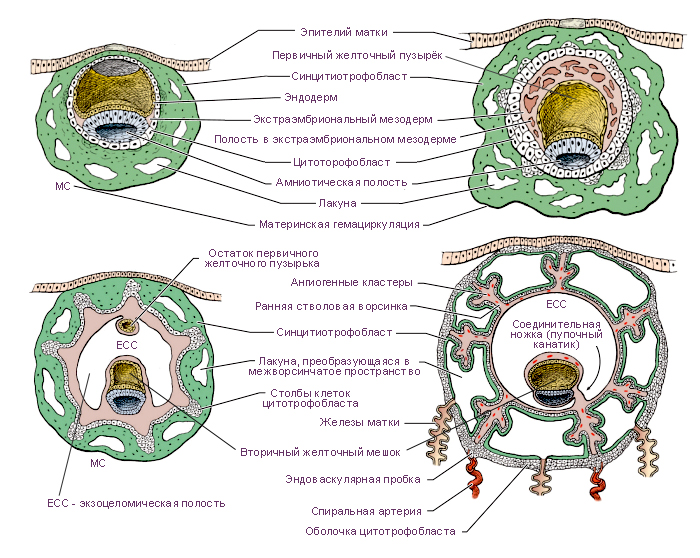
Во время первой фазы гаструляции эмбриобласт расслаивается на эпибласт – слой цилиндрических клеток, и гипобласт – слой кубических клеток, обращенных к бластоцелю. Эпибласт и гипобласт вместе образуют двухслойный (биламинарный) зародышевый диск (бластодиск). В это же время между клетками эпибласта появляется небольшая полость, которая постепенно увеличивается в размерах и становится амниотической полостью.
Клетки гипобласта выселяются из внутренней клеточной массы вследствие слабого адгезионного взаимодействия между ними. Интенсивно пролиферирующие клетки гипобласта перемещаются по внутренней поверхности цитотрофобласта и формируют выстилку экзоцеломической полости, или первичного желточного мешка. Этот слой будет называться внезародышевой энтодермой.
Вскоре после этого между внутренней поверхностью цитотрофобласта и наружной поверхностью экзоцеломической полости появляется новая популяция клеток, которая образует рыхлую внезародышевую мезодерму. В этом слое образуются увеличивающиеся в размерах полости, которые постепенно сливаются и формируют новую внезародышевую хорионическую полость.
Во время второй фазы гаструляции краниальный (головной) отдел зародышевого диска расширяется, а в суженной каудальной (хвостовой) части зародышевого диска на 15-е сутки возникает вытянутое в кранио-каудальном направлении утолщение эпибласта – первичная полоска. На переднем конце первичной полоски образуется скопление клеток — Гензеновский, или первичный, узелок. Через него мигрируют клетки, и образуются энтодерма, мезодерма, эктодерма и хорда.
Мигрирующие клетки подворачиваются вниз и перемещаются в краниальном направлении. Часть мигрирующих клеток Гензеновского узелка и передней трети первичной полоски встраивается в гипобласт и принимает участие в формировании прехордальной пластинки и зародышевой энтодермы. Остальной материал первичной полоски перемещаются в пространство между эпибластом и гипобластом, и образует зародышевую мезодерму. Клетки, которые остаются в эпибласте, образуют зародышевую эктодерму, содержащую эмбриональные зачатки кожной эктодермы и нервной трубки.
Хорда зародыша – это своеобразный клеточный стержень, который определяет первичную ось эмбриона. Хорда развивается из нотохорда (головного отростка). Хорда формирует ось скелета зародыша и является основой развития костей осевого скелета (позвоночника, ребер, грудины, черепа). Вокруг хорды в будущем сформируется позвоночный столб. Остатки хорды сохраняются в виде студенистого ядра межпозвоночного диска.
НЕЙРУЛЯЦИЯ
В результате гаструляции формируются зародышевые листки, части которых, влияя друг на друга, индуцируют образование новых структур. Пример такого влияния – дифференцировка прилежащего к нотохорду сверху участка эктодермы в нервную пластинку. Нейруляция – процесс закладки и развития нервной трубки. Нейруляция начинается с 16-х суток развития и, в основном, завершается к 23-м.
Стадии нейруляции
первичная эмбриональная индукция
(процесс образования нервной пластинки из дорсальной эктодермы);
приподнимание краев нервной пластинки и образование нервного желобка;
появление нервных валиков (возвышений нервной пластинки по обе стороны от желобка);
смыкание нервных валиков по срединной линии с образованием замкнутой нервной трубки;
срастание эктодермы над нервной трубкой;
формирование нервного гребня и начало выселения из него клеток.
Краниальный и каудальный участки нервной трубки долго остаются незамкнутыми, их называют соответственно передним и задним нейропором. Передний нейропор закрывается на 23-26-й день развития, а задний – на 26-30-й день. После смыкания обоих нейропоров, образуется замкнутая трубчатая структура – центральная нервная система, более узкая в каудальной части (спинной мозг) и расширенная в краниальном отделе (головной мозг).
После смыкания валиков и образования нервной трубки часть эктодермы, расположенная между нейральной и кожной эктодермой, формирует новую структуру – нервный гребень. Нервный гребень дает начало периферической нервной системе, а также одонтобластам, меланоцитам, Шванновским клеткам, клеткам мозгового вещества надпочечников и др.
Нейрогенные плакоды – это утолщения эктодермы, расположенные латерально по обе стороны от формирующейся нервной трубки в краниальном отделе зародыша. Из нейрогенных плакод формируется хрусталик глаза, слуховой пузырёк, обонятельный эпителий.
ДЕЦИДУАЛЬНАЯ РЕАКЦИЯ
Изменения эндометрия при имплантации были названы децидуальной реакцией, а эндометрий беременной матки – децидуальная (отпадающая) оболочка. Во время этой реакции эндометрий утолщается и его стромальные клетки, увеличиваясь, становятся децидуальными клетками. Они содержат много липидов и гранул гликогена, секретируют пролактин и простагландины. Децидуальная реакция предотвращает инвазию трофобласта глубже эндометрия. Различают три части децидуальной оболочки.
Базальная децидуальная оболочка (decidua basalis) расположена между эмбрионом (затем плодом) и стенкой матки, т.е. в месте имплантации, она формирует материнскую часть плаценты.
Капсулярная децидуальная оболочка (decidua capsularis) покрывает имплантированный эмбрион (затем плод) и отделяет его от полости матки. По мере развития плода капсулярная часть децидуальной оболочки растягивается и дегенерирует.
Париетальная децидуальная оболочка (decidua parietalis) представляет собой остатки эндометрия на противоположной стороне матки, т.е. часть, которая напрямую не контактирует с эмбрионом.
ПРОВИЗОРНЫЕ ОРГАНЫ
Часть бластомеров и клеток после дробления зиготы идет на образование временных органов, способствующих развитию зародыша и плода. Они образуют оболочки зародыша, связывают его с организмом матери и выполняют некоторые специальные функции. Такие органы называются провизорными, или внезародышевыми.
У человека, развиваются четыре внезародышевых органа:
-
хорион;
-
амнион;
-
желточный мешок;
аллантоис.
Хорион
Хорион (ворсинчатая оболочка) – это провизорный орган зародыша, развивающийся из трофобласта и подстилающей его мезодермы; хорион образует выросты (ворсины), врастающие в слизистую оболочку матки и образующие плодовую часть плаценты.
В процессе имплантации трофобласт образует на наружной поверхности первичные ворсины, являющиеся выпячиванием синцитиотрофобласта и цитотрофобласта (Рис. 7). На 12-13-й день при прорастании подлежащей внезародышевой мезодермы в первичные ворсины, они превращаются во вторичные ворсины, и соответственно состоят (снаружи внутрь) из синцитиотрофобласта, цитотрофобласта и стержня из внезародышевой мезодермы.
Однако постепенно (начиная с 3-й недели развития) на большей части хориона ворсинки редуцируются и сохраняются только в той части хориона, которая направлена к базальной децидуальной оболочке. Этот период называют плацентацией. При этом ворсинки разрастаются, в них врастают сосуды, которые позже соединятся с пупочными сосудами плода. Третичные ворсины состоят из синцитиотрофобласта, цитотрофобласта, внезародышевой мезодермы и кровеносных сосудов. Позже цитотрофобласт исчезает, так как все его клетки сливаются с синцитиотрофобластом.
При развитии хориона выделяют два периода:
1) период ворсинчатого хориона;
2) период гладкого хориона.
В период ворсинчатого хориона (chorion frondosum) последовательно образуются первичные, вторичные и третичные ворсины. Ворсины равномерно распределены по всей поверхности эмбриона. Но не все ворсины хориона развиты одинаково хорошо. Ворсины, обращенные к капсулярной части децидуальной оболочки, развиты слабо и постепенно исчезают. Поэтому хорион в этой части называют гладким (chorion laeve).
Третичные ворсины, прилежащие к базальной децидуальной оболочке образуют плодовую часть плаценты.
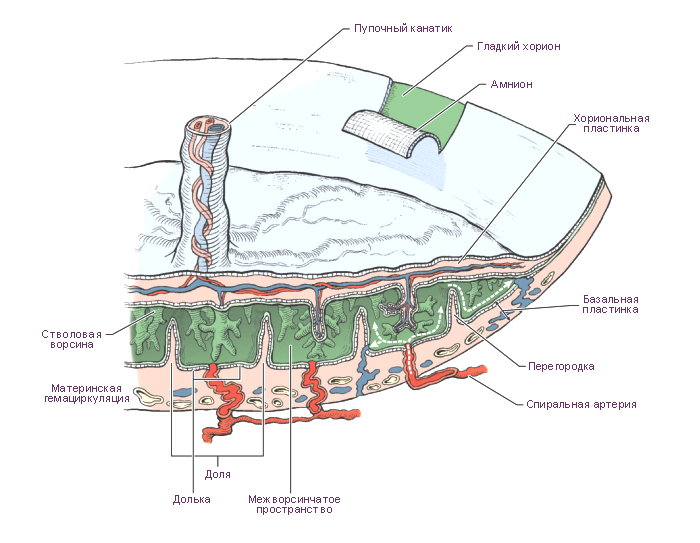
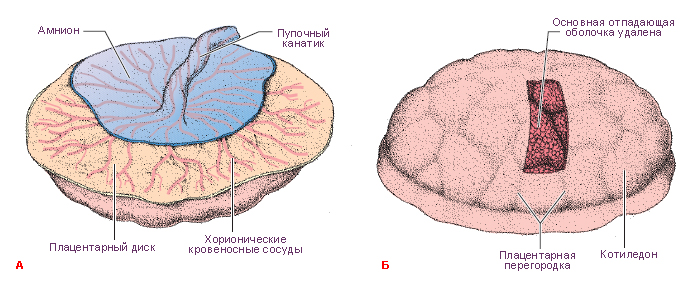
Хорионическая полость в это время становится больше, и к 19-20-му дню эмбрион соединяется с трофобластом посредством узкого плодного стебелька (соединяющего стебелька). Плодный стебелек позже образует пупочный канатик (пуповину), который соединяет зародыш, а затем и плод, с плацентой.
Функции хориона: защитная, трофическая, газообменная, экскреторная и другие, в которых хорион принимает участие, будучи составной частью плаценты.
Амнион
Амнион – плодный пузырь – объемистый мешок, заполненный амниотической жидкостью (околоплодными водами). В области пупочного кольца амнион переходит на пуповину и далее на плодовую часть плаценты.
Клетки амниона продуцируют амниотическую жидкость (околоплодные воды), в которой развивается эмбрион, а затем – плод.
Одновременно с расслоением внутренней клеточной массы на эпибласт и гипобласт образуется амниотическая полость, ограниченная эпибластом и внезародышевой (амниотической) эктодермой. В ходе гаструляции клетки внезародышевой мезодермы обрастают амниотическую эктодерму, формируя наружный слой амниона.
Стенка амниона состоит из:
внезародышевой эктодермы;
внезародышевой мезодермы.
Функции амниона — образование околоплодных вод и защитная функция. Амниотическая жидкость защищает зародыш при сотрясении, позволяет плоду совершать движения и предотвращает слипание растущих частей тела друг с другом и с окружающими тканями.
После рождения ребенка хорион и амниотическая оболочка отторгаются.
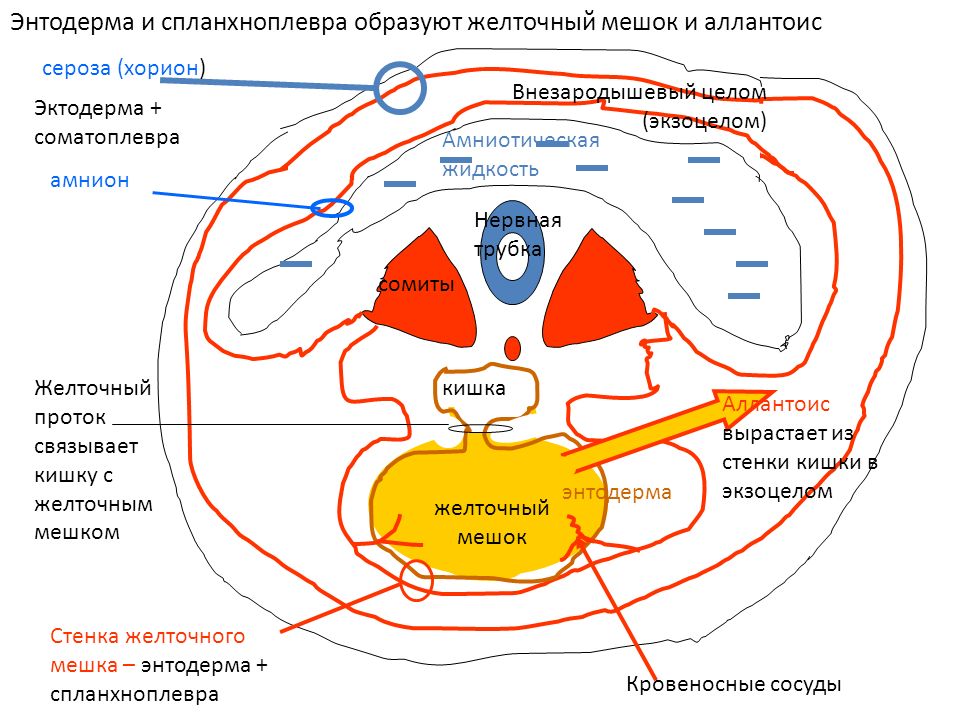
Желточный мешок
Желточный мешок – вынесенная за пределы зародыша часть первичной кишки.
Стенка желточного мешка состоит из: 1) внезародышевой энтодермы;
2) внезародышевой мезодермы.
По мере развития амнион сдавливает желточный мешок; образуется узкая перемычка, соединяющая его с полостью первичной кишки, — желточный стебелек. Эта структура удлиняется и вступает в контакт с плодным стебельком, содержащим аллантоис. Желточный стебелек и дистальная часть аллантоиса вместе со своими сосудами образуют пупочный канатик, отходящий от зародыша в области пупочного кольца.
Желточный мешок выполняет следующие функции:
1) кроветворную (в стенке желточного мешка (в мезодерме) образуются первые клетки крови, которые затем мигрируют в кроветворные органы зародыша);
2) гонобластическую (в стенке желточного мешка (в энтодерме) образуются первичные половые клетки (гонобласты), которые затем мигрируют в закладки половых желез зародыша).
Аллантоис
Аллантоис – слепое выпячивание передней стенки задней кишки, прорастающее в плодный стебелек (будущий пупочный канатик). Формирование аллантоиса начинается на 16-й день эмбриогенеза. Аллантоис образован зародышевой энтодермой.
У животных, развивающихся в яйце, аллантоис достигает большого развития и выполняет функцию резервуара для продуктов обмена зародыша (главным образом мочевины). Именно поэтому аллантоис нередко называют мочевым мешком.
У млекопитающих необходимость в накоплении продуктов обмена отсутствует, так как они поступают через плацентарный кровоток в организм матери и выводятся ее экскреторными органами. Поэтому у таких животных и человека аллантоис развит слабо и имеет важное значение в эмбриональном кроветворении и ангиогенезе: в его стенке развиваются пупочные сосуды, которые разветвляются в плаценте и благодаря которым формируется плацентарный круг кровообращения. Плодный стебелек вместе с сосудами вытягивается и превращается в пупочный канатик, а сосуды (две артерии и вена) называются пупочными сосудами.
Функция аллантоиса — способствование выполнению функций плаценты.
На 2-м месяце эмбриогенеза аллантоис дегенерирует, а на его месте появляется урахус – плотный фиброзный тяж, тянущийся от верхушки мочевого пузыря к пупочному кольцу. В постнатальном периоде урахус организуется в срединную пупочную связку.
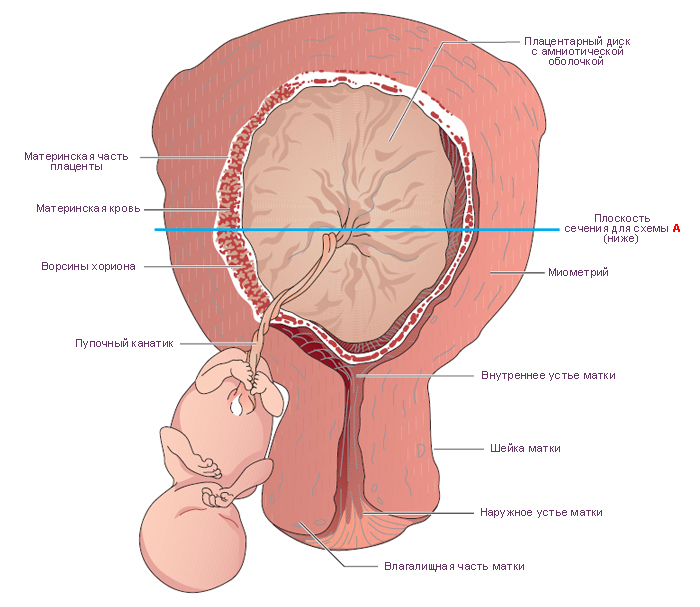
ПЛАЦЕНТА
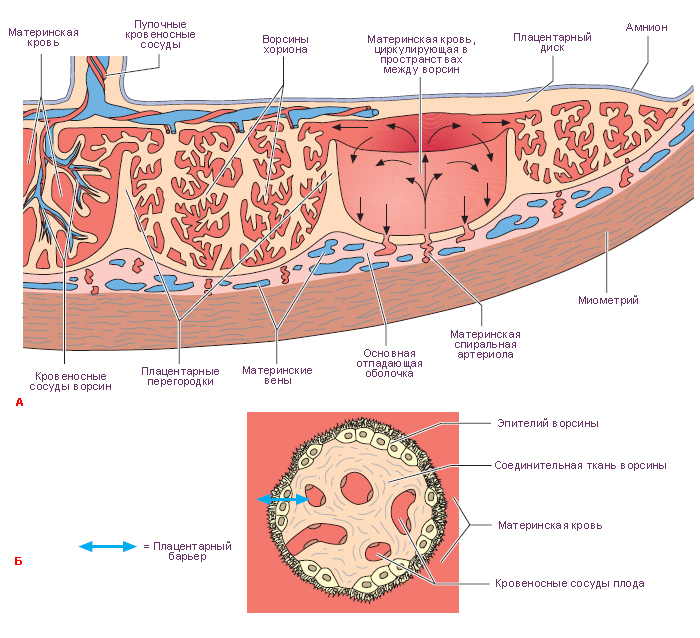
Плацента — это временный орган, который начинает формироваться во время имплантации. Плацента связывает плод с организмом матери и состоит из плодового (ворсинчатый хорион) и материнского (базальная децидуальная оболочка) компонентов. Она транспортирует питательные вещества и кислород от матери к эмбриону, очищает кровь плода от токсинов и углекислого газа и секретирует гормоны.
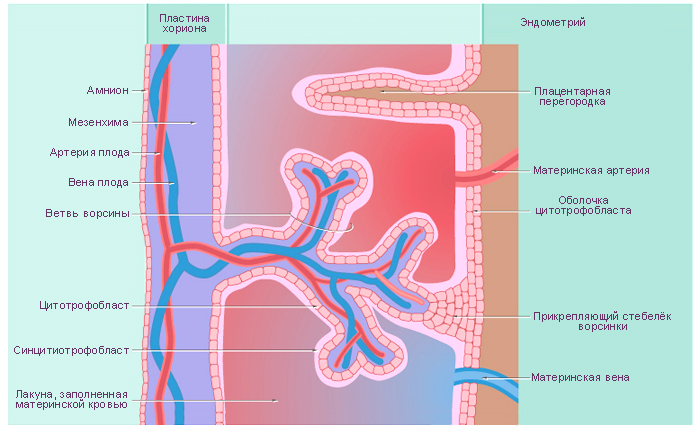
Стадии развития плаценты (плацентация). Погружающийся в стенку матки синцитиотрофобласт окружает маленькие островки эндометрия, содержащие кровеносные сосуды. Ферменты синцитиотрофобласта лизируют (растворяют) материнскую ткань, оставляя пространства, или лакуны, и разрывая кровеносные сосуды. Кровь из поврежденных сосудов заполняет лакуны материнской кровью. Ворсины хориона прорастают в эти лакуны и развиваются для того, чтобы, во-первых, кровь в сосудах плода и материнская кровь в лакунах находились достаточно близко для полноценного обмена, во-вторых, для формирования избирательно проницаемого плацентарного барьера.
Структурно-функциональной единицей сформировавшейся плаценты является котиледон. Он образован стволовой ворсиной и ее разветвлениями, содержащими сосуды плода. Стволовая или якорная, ворсина представляет собой большой вырост хориона, плотно сросшийся с базальной децидуальной оболочкой, от боковой поверхности которого отходят многочисленные разветвления мелких ворсин хориона. К 140-му дню беременности в плаценте сформировано около 10-12 больших, 40-50 мелких и до 150 рудиментарных котиледонов. Соседние котиледоны разграничены соединительнотканными перегородками – септами, по которым проходят артериальные сосуды, несущие обогащенную кислородом и питательными веществами кровь к лакунам плаценты. В лакуны широкими отверстиями открываются лакунарные вены, по которым материнская кровь оттекает из плаценты. Стенка лакун образована соединительной тканью эндометрия, которая покрыта наслоениями аморфной субстанции, так называемым фибриноидом.
Фибриноид – неклеточная фибриноподобная масса, которая появляется на поверхности ворсин со второй половине беременности. По-видимому, она представляет собой смешанный продукт распада эпителия ворсин и свёртывания плазмы материнской крови.
К 4-му месяцу беременности формирование основных структур плаценты заканчивается. Лакуны полностью сформированной плаценты содержат около 150 мл материнской крови, полностью обменивающейся в течение 3—4 мин.
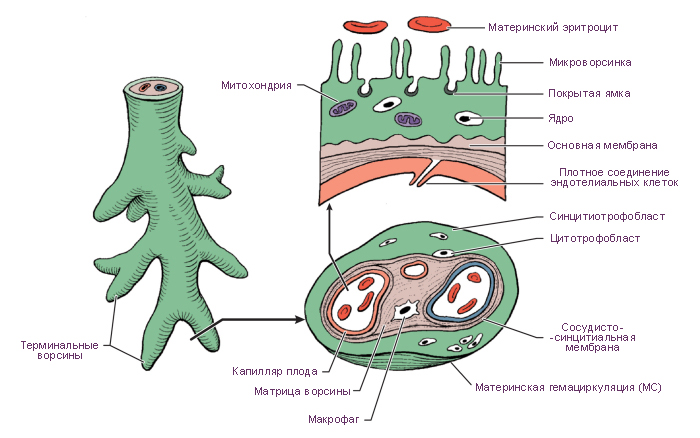
Плацентарный барьер
К 23-му дню гестации кровь эмбриона циркулирует в третичных ворсинах. Питательные вещества из материнской крови в лакунах достигают эмбриона, а позднее – плода, последовательно проходя через:
синцитиотрофобласт;
цитотрофобласт (который позже исчезает);
базальную пластинку трофобласта;
внезародышевую мезодерму;
базальную пластинку сосудов третичных ворсин ;
эндотелиальные клетки сосудов плода.
Эти 6 слоев образуют плацентарный барьер, который отграничивает проникновение ряда веществ из материнского кровотока в плодовый.
Функции плаценты
Функции плаценты многообразны. К ним относятся метаболический обмен между матерью и плодом, транспорт материнских антител, эндокринная функция (синтез хорионического гонадотропина человека, прогестерона, хорионического соматоммаммотропина, трасферрина, пролактина), детоксикация метаболитов, лекарственных препаратов.
ПРОИЗВОДНЫЕ ЗАРОДЫШЕВЫХ ЛИСТКОВ
Производные эктодермы
Наружный зародышевый листок дифференцируется на:
нейроэктодерму (из нее развивается нервная трубка и нервный гребень);
кожную эктодерму (развивается эпидермис и производные кожи).
Производные энтодермы
Из внутреннего зародышевого листка формируется эпителиальная выстилка первичной кишки и дыхательных путей, а также паренхима щитовидной и паращитовидной желез, печени и поджелудочной железы.
Производные мезодермы
Клетки зародышевой мезодермы выселяются из эпибласта и формируют дорсальную мезодерму, из которой возникают сомиты – симметричные парные структуры по бокам от хорды и нервной трубки. Из мезодермы образуется еще два зачатка: нефротом (промежуточная мезодерма) и латеральная мезодерма.
Дорсальная мезодерма. В каждом сомите различают склетором, дерматом и миотом; их клетки имеют свои пути миграции и служат источником для различных структур.
Клетки склеротома интенсивно размножаются и выселяются из сомита, окружая хорду и вентральную (переднюю) часть нервной трубки. Выселившиеся клетки дифференцируются в хрящевые и образуют позвонки, ребра и лопатки.
В оставшейся части сомита выделяют миотом (внутренний слой клеток, образующий впоследствии скелетную мускулатуру) и дерматом (наружный слой, зачаток соединительнотканной части кожи).
Промежуточная мезодерма (нефротом). Латеральнее сомитов находится скопление клеток промежуточной мезодермы (нефротом) – зачаток мочевой и половой систем.
Латеральная мезодерма. Расположенная латеральнее нефротома мезодерма расщеплена на два листка: соматическая мезодерма, из нее образуются конечности и стенки тела (например, париетальная серозная оболочка), и спланхническая мезодерма, из которой образуются сердце, кора надпочечников, строма гонад, соединительная и гладкомышечная ткани внутренних органов и кровеносных сосудов.
Пингбэк: Половая система — yavmede.ru