


Свет – электромагнитная волна со скоростью распространения в вакууме 300 000 км/сек. Кроме того, он имеет и свойства частиц (фотонов), которые не имеют массы покоя и заряда, но обладают энергией. Таким образом, свет имеет корпускулярно-волновую природу.
Цвет – одна их характеристик света. Цвет определяется длинной волны. Видимый свет для глаза человека имеет длину волны от 380 нм (фиолетовый) до 760 (красный). Энергия света зависит от длины волны (обратно пропорциональна, т.е. чем больше длина волны, тем меньше энергия луча). От энергии светового луча зависит глубина его проникновения в биологический объект и возникающие при этом реакции
(фотохимические реакции, фототоксический эффект и т.д.).
При прохождении света через объект часть его энергии поглощается. Для характеристики поглощающей способности среды используют понятие коэффициент поглощения света. Он зависит от длины волны и свойств среды.
При переходе луча из одной среды в другую он меняет направление (преломляется), поскольку в разных средах свет распространяется с разной скоростью. Чем больше разница скоростей луча в разных средах – тем сильнее он преломляется.
Микроскопы

- прямой (объектив находится над препаратом);
- инвертированный (инвертор; объектив находится под препаратом; применяется при изучении живых клеток в культуре).
Источник света
Дневной свет или лампа накаливания (галогеновая лампа дает яркое постоянное освещение). Во флуоресцентной микроскопии применяют ртутные или ксеноновые лампы, лазеры с различной длинной волны.
Коллекторная линза – для настройки освещения образца.
Полевая диафрагма – для изменения площади освещения образца (не влияет на разрешение изображения).
Конденсор
Система линз, собирающих лучи от источника света таким образом, чтобы освещение объекта было равномерным.
Ирисовая (апертурная) диафрагма: позволяет регулировать размеры апертуры (отверстия), через которое свет идет к объекту (это влияет на контраст, глубину фокуса и разрешение изображения). Установлена в конденсоре.
Объектив
Собирает лучи света от объекта и создает первичное перевернутое истинное изображение объекта. Несколько объективов установлены в револьверной головке. При смене объективов фокусное расстояние сохраняется (объект остается в фокусе).
Окуляр
Переводит первичное действительное изображение в мнимое вторичное. Корректирует хроматические аберрации.
Увеличение микроскопа определяется произведением увеличений объектива и окуляра.
Техника микроскопирования и правила работы с микроскопом.
Установить микроскоп у края стола против левого плеча. С правой стороны расположить открытый альбом для практических занятий. Протереть окуляр марлевой салфеткой. Обратить зеркало микроскопа вогнутой стороной к источнику света.
Привести микроскоп в рабочее состояние:
установить объектив малого увеличения на расстоянии 1-1,5 см от предметного столика;
поднять конденсор до предела;
смотря в окуляр одним глазом, не закрывая другого, равномерно и интенсивно осветить зеркалом поле зрения;
Поместить препарат на предметный столик микроскопа покровным стеклом кверху (обратить на это внимание). Передвигать препарат большим и указательным пальцами за ребро с тем, чтобы найти место, подлежащее изучению; средний палец должен упираться в предметный столик.
Опустить конденсор до положения, обеспечивающего наилучшее освещение препарата на малом увеличении.
Рассмотреть препарат с помощью 8- или 10-кратного объектива (малого увеличения). Найти место, подходящее для изучения при большом увеличении, поставить его в центр поля зрения и отрегулировать резкость изображения микровинтом.
При переходе с малого увеличения на большое необходимо, не меняя фокусного расстояния, повернуть револьверную пластинку объективов до щелчка, чтобы в рабочее положение встал 40-кратный объектив (большого увеличения). Микровинтом добиться резкого изображения. Работа макровинтом на большом увеличении запрещается, так как легко можно раздавить препарат.
Левая рука должна находиться на микрометрическом винте, слегка поворачивая его в обе стороны для просматривания деталей среза, лежащих на разной глубине поверхности среза.
Уяснив пространственное положение, пропорции и взаимоотношения деталей изучаемого объекта, приступить к зарисовке.
Запрещается снимать препарат из под объектива большого увеличения. Для того, чтобы убрать препарат с предметного столика микроскопа в рабочее положение ставится объектив малого увеличения и препарат снимается с предметного столика микроскопа.
Запрещается развинчивать какие-либо части микроскопа. В случае неисправности микроскопа обращаться к преподавателю
Типы микроскопии
Светлое поле
Используется чаще всего. Препарат должен быть прозрачен и окрашен
Темное поле
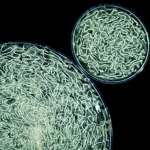
Темнопольный микроскоп применяется для получения изображений прозрачных живых объектов. Используется эффект Тиндаля (который позволяет видеть пылинки в луче света в темной комнате).
Под конденсором установлена заглушка таким образом, чтобы свет попадал только на края конденсорной линзы. Поэтому свет, выходящий из конденсора, не попадает в объектив: при отсутствии образца видно только темное поле.
При наличии препарата свет, проходя через него, отклоняется и попадает в объектив: объект выглядит очень светлым на темном фоне (с очень большим контрастом). Метод предъявляет высокие требования к чистоте препарата.
Фазовый контраст
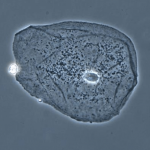
Фазово-контрастный микроскоп применяется для исследования изображений прозрачных (неокрашенных) объектов, особенно живых клеток в культуре. Препарат должен быть тонким (монослой клеток). В отличие от темнопольной микроскопии, метод позволяет оценить не только контуры объекта, но и его структуру. Высокое разрешение получить трудно.
Принципы фазово-контрастной микроскопии сформулированы голландским физиком Фрицем Цернике в 1934 г. В основе метода лежит следующее явление. Скорость света зависит от среды, через которую он проходит. При переходе из воздуха в более плотную среду электромагнитная волна замедляется. Поскольку разные участки препарата имеют разные показатели преломления, то лучи, прошедшие через оптически более плотные участки, будут отставать от лучей, прошедших через оптически менее плотные участки – возникает фазовый сдвиг между лучами. Величина сдвига зависит от показателей преломления сред и длины пути, пройденного в каждой среде. Однако фазовый сдвиг не улавливается глазом человека. Поэтому специальные устройства (заглушка; фазовая пластинка в объективе) преобразуют фазовый сдвиг в различия в интенсивности (яркости) света, которые воспринимаются глазом.
Фриц Цернике (нидерл. Frits Zernike; 16 июля 1888-10 марта 1966) — голландский физик, лауреат Нобелевской премии по физике 1953 года «За обоснование фазово-контрастного метода, особенно за изобретение фазово-контрастного микроскопа».
Поляризационная микроскопия
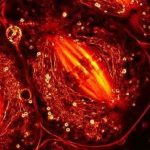
Свет – ЭМ волны, колебания которых происходят во всех плоскостях перпендикулярно направлению распространения света. Существуют материалы, которые пропускают ЭМ волну, колебания которой происходят строго в определенной плоскости. Такие материалы называют анизотропными, из них делают поляризаторы. После прохождения света через поляризатор он становится поляризованным (колебания происходят в одной плоскости). Если на пути такого луча поставить второй поляризатор (анализатор), повернутый вокруг вертикальной оси под прямым углом к первому, то свет через него проходить не будет. На этом свойстве основана поляризационная микроскопия.
Свет проходит через поляризующий фильтр и полностью «запирается» анализатором. Однако если на пути света поместить препараты, способные менять плоскость поляризации, образуются световые волны, которые проходят через анализатор: на темном фоне возникает яркое изображение объекта. Структуры, способные
менять плоскость поляризации (т.е. обладающие анизотропными свойствами): волокна коллагена, миофибриллы, нейрофибриллы, кристаллы.
Интерференционная микроскопия

Интерференционная микроскопия – это дальнейшее развитие фазово-контрастной и поляризационной микроскопии. Как и фазово-контрастная микроскопия, этот метод дает возможность наблюдать прозрачные и бесцветные объекты, но их изображения могут быть и разноцветными (интерференционные цвета). Оба метода пригодны для изучения живых тканей и клеток и применяются во многих случаях именно с этой целью.
Суть метода состоит в том, что каждый луч, входящий в микроскоп, раздваивается. Один из полученных лучей направляется сквозь наблюдаемую частицу, другой — мимо неё. В окулярной части микроскопа оба луча вновь соединяются и интерферируют между собой. Результат интерференции определяется разностью хода лучей (луч, проходящий через объект, запаздывает по фазе). Величина этого запаздывания измеряется компенсатором. Можно сказать, что метод интерференционного контраста сходен с методом фазового контраста — они оба основаны на интерференции лучей, прошедших через микрочастицу и миновавших её. Главное отличие интерференционной микроскопии от метода фазового контраста – это возможность измерять разности хода, вносимые микрообъектами.
В методе дифференциального интерференционного контраста (ДИК) обе волны проходят через один и тот же объект с небольшим боковым смещением. Наибольшее распространение получил вариант ДИК по Номарскому, в котором разделение и сведение пучков производятся в поляризованном свете с помощью специальных двоякопреломляющих призм, установленных соответственно перед конденсором и после объектива. Величина разведения пучков выбирается близкой к разрешающей способности микроскопа, чтобы не было заметно двоение изображения. Получаемое цветное изображение рельефно.
Люминисцентная микроскопия
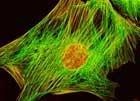
Люминесцентный микроскоп оснащен специальным источником света, испускающим ультрафиолетовый или синий свет. Поглощая это излучение, образец испускает свет большей длины волны – видимый свет люминесценции
(аутофлуоресценция). Также используется для выявления элементов, меченых флуоресцентными метками (флуорохромами). Флуорохромы также при освещении испускают свет большей длины волны (видимой части спектра).
Фильтр между источником света и препаратом отсекает все лучи, кроме тех, необходимых для возбуждения флуоресценции. Специальный фильтр между объективом и окуляром защищает глаза от ультрафиолета и пропускает только испускаемый препаратом видимый свет. Люминесцентные микроскопы широко используются в биологии, а также в медицине для диагностики.
Электронная микроскопия
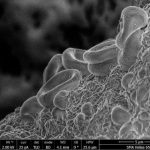
Вместо света используется пучок электронов (длина волны – 0,005 нм), что позволяет увеличить разрешение и увеличение в 200 раз и более. Стеклянные линзы не прозрачны для электронов, но пучок электронов может отклоняться и фокусироваться ЭМ полем.
Аналогами источника света являются катод и анод. Катод(-) – металлическая нить, которая испускает электроны, которые нагреваются в вакууме. Анод(+) – пластинка с отверстием в центре. Разница зарядов катода и анода ускоряет электроны, часть из них проходит через отверстие в аноде и формирует пучок электронов. Конденсором является ЭМ поле, которое фокусирует пучок на образце. Образец – ультратонкий срез, окрашенный (контрастированный) с использованием электроно- поглощающего или рассеивающего вещества.
Формируемое изображение – тень контрастного материала (электроно-плотных компонентов образца). ЭМ объектив отклоняет часть пучка, которая прошла через образец, чтобы сформировать и увеличить изображение. Далее 1 или 2 ЭМ
проектора (аналоги окуляра) еще увеличивают изображение и проецируют его на фотоэмульсию или флуоресцентный экран.

ТЕМ (трансмиссионная, или просвечивающая, электронная микроскопия) – позволяет увидеть внутреннюю структуру клеток и тканей (предел разрешения – 0,2 нм). Контрастирующие вещества выбирают по способности поглощать электроны и аффиности к конкретным клеточным компонентам. Используют соли тяжёлых металлов, например, цитрат свинца. Четырёхокись осмия взаимодействует с липидпми и применяется для окрашивания клеточных мембран.
SEM (сканирующая электронная микроскопия) – позволяет увидеть поверхностные ультраструктурные элементы (предел разрешения – 2 нм). Образцы не окрашивают. Сначала образец высушивают путем перехода критической точки. Для этого после дегидратации образцы вымачивают в растворах, смешивающихся с СО2 или фреоном, а затем помещают в камеру критического высушивания, температура в которой составляет 31градус, а давление 73 атм. В этих условиях изменяется поверхностное натяжение и жидкость беспрепятственно покидает образец, не нарушая его структуры. Затем образец покрывают тонким слоем тяжелого металла, узкий пучок электронов направляется перпендикулярно к поверхности образца, последовательно, точка за точкой, и генерируются 2 сигнала:
- Вторичные электроны, освобожденные с поверхности образца, собираются на детекторе и конвертируются в изображение на катодной лучевой трубке.
- Рентгеновские лучи генерируются, когда пучок выбивает атомы тяжелее натрия (анализ этих сигналов может дать информацию о концентрации и распределении некоторых элементов в образце).
Принципиально новые возможности электронной микроскопии открылись сравнительно недавно, после разработки метода «замораживания – скалывания». С помощью этого метода исследуются тончайшие детали строения клетки, при этом получается объемное изображение в трансмиссионном электронном микроскопе.
При обычном замораживании в клетках образуются кристаллики льда, которые заметно искажают их структуру. Во избежание этого клетки замораживают очень быстро при температуре жидкого азота (- 196 С). При таком мгновенном замораживании кристаллы льда не успевают образоваться, и клетка не испытывает деформаций.
Замороженный блок раскалывают лезвием ножа (отсюда и название метода). Затем, обычно в вакуумной камере, избыток льда удаляют возгонкой. Эта операция называется травлением. После травления более резко обозначается рельеф в плоскости скола. Полученный образец оттеняется, то есть на поверхность образца напыляется тонкий слой тяжелых металлов. Однако весь фокус состоит в том, что напыление производится под углом к поверхности образца. Это очень важный момент. Появляется эффект тени, изображение выглядит объемным.
В трансмиссионном микроскопе электронный луч способен проникнуть только через очень тонкие срезы. Обычная толщина оттененных образцов чрезмерно велика, поэтому органическую материю, подстилающую слой металла, необходимо растворить. В результате остается тонкая металлическая реплика (или отпечаток) с поверхности образца. Реплику и используют в трансмиссионном микроскопе.
Этот метод предоставил, например, уникальную возможность наблюдать внутреннее строение мембран клетки.
Общая характеристика
Клетка – это структурная и функциональная единица живого во всех органах и тканях. Существует два типа клеток: прокариоты – мелкие одноклеточные организмы, которые утратили ядерную оболочку, гистоны и мембранные органеллы.
Эукариотическая клетка существует в первую очередь как компонент многоклеточного организма.
Компоненты эукариотической клетки:
1. Клеточная мембрана отделяет клетку от ее окружения и формирует различные функциональные зоны внутри клетки (ядро, органеллы). Внешняя мембрана клетки называется плазматическая мембрана, или плазмолемма.
2. Цитоплазма окружает ядро и находится внутри плазмолеммы. Она содержит структуры и вещества, необходимые для реализации программы, заложенной в ДНК, и выполнения клеткой своих функций.
3. Ядро клетки содержит ДНК, в которой закодирована генетическая информация, необходимая для синтеза белков и жизнедеятельности клетки. Оно также содержит компоненты, которые помогают определить, какая часть генетического кода будет использоваться, и которые передают эту информацию в цитоплазму.
Функции клетки:
• Рост
• Развитие
• Воспроизводство
КЛЕТОЧНАЯ МЕМБРАНА
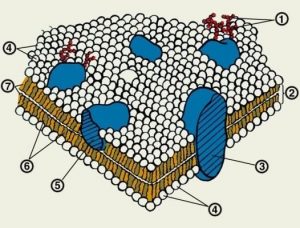
1. Липиды в клеточной мембране представлены фосфолипидами, сфинголипидами и холестеролом. Больше всего в мембране фосфолипидов. Каждая молекула фосфолипида имеет гидрофильную головку, содержащую фосфаты, и два гидрофобных хвоста из жирных кислот. Фосфолипиды образуют двойной слой (бислой) таким образом, чтобы хвосты были направлены друг к другу, в центр мембраны. Слой мембраны, обращенный к цитоплазме, называется Р-поверхность (протоплазматическая), а слой, обращенный наружу – E-поверхность
(эктоплазматическая).
2. Белки составляют более половины веса мембраны. Большинство белков мембраны глобулярные, и принадлежат к одной из двух групп:
• Интегральные мембранные белки погружены в липидный бислой глубоко, для их выделения необходимы детергенты. Некоторые из них выступают над мембраной только с одной стороны, другие прободают ее полностью и выступают над поверхностью мембраны с обеих сторон (трансмембранные белки). Гидрофильные аминокислоты этих белков контактируют с фосфатными группами фосфолипидов, а гидрофобные аминокислоты – с гидрофобными хвостами. Некоторые трансмембранные белки могут образовывать гидрофильные каналы, пропускающие воду и водорастворимые компоненты через гидрофобную часть мембраны. Интегральные белки могут подвергаться реорганизации благодаря ассоциации с периферическими белками, филаментами цитоскелета, мембранами соседних клеток, компонентами внеклеточного матрикса.
• Периферические мембранные белки прилежат к внутренней или наружной стороне мембраны и могут быть как глобулярными, так и фибриллярными. Например, у эритроцитов на Р-поверхности расположен спектрин, который поддерживает целостность мембраны, и анкирин, который связывает спектрин с трансмембранными белками.
3. Углеводы присутствуют в мембране главным образом в виде олигосахаридных остатков мембранных гликопротеинов и гликолипидов. Мембранные олигосахариды выступают над наружной поверхностью мембраны, формируя гликокаликс, который участвует в клеточной адгезии и распознавании.
Функции мембраны
1. Избирательная (селективная) проницаемость. Клеточная мембрана формирует эффективный барьер между клеткой или органеллами и их внешним окружением, что предотвращает попадание вредных веществ внутрь и размывание макромолекул, ферментов и субстратов. Селективная проницаемость мембраны имеет важное значение для поддержания постоянного состояния среды (гомеостаза), необходимого для выживания клетки. Этот механизмы обеспечивает оптимальную концентрацию ионов, воды, ферментов и субстратов внутри клетки.
• Пассивная диффузия. Некоторые вещества, например, вода, могут проходить через мембрану напрямую, по градиенту концентрации. Этот процесс не связан с затратами энергии.
• Облегченная диффузия. Некоторые вещества, например, глюкоза, могут проходить через мембрану с помощью ее компонентов. Это процесс часто происходит только в одном направлении, требует наличия градиента концентрации и не требует затрат энергии.
• Активный транспорт. Некоторые вещества могут проходить через мембрану в обоих направлениях независимо от градиента концентрации. Этот процесс требует затрат энергии, обычно в виде АТФ. Примером такого транспорта является калий-натриевый насос, который позволяет выводить натрий из клетки даже если его концентрация вне клетки выше, чем внутри.
2. Передача сигнала (сигнальная трансдукция). На поверхности клетки расположены рецепторы, которые специфично воспринимают экзогенные сигналы (например, гормональные). Сигнальная молекула, которая связывается с рецептором, называется лигандом. После того, как лиганды свяжутся с рецепторами, они могут передать сигнал внутрь клетки следующими путями:
• Через ассоциацию с компонентами цитоскелета на внутренней поверхности клетки.
• Через взаимодействие с другими компонентами мембраны, что приводит к продукции молекулы вторичного посредника (мессенджера), который затем несет сигнал внутрь клетки.
• Рецептор сам может быть ферментом. Связывание лиганда с рецептором активирует его ферментативную активность, он расщепляет внутриклеточные белки, передавая таким образом сигнал в клетку.
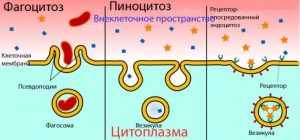
3. Эндоцитоз. Клетки поглощают внеклеточные вещества, и они поступают в цитоплазму в ограниченных мембраной пузырьках посредством эндоцитоза.
• Фагоцитоз («клеточная еда») – клетки поглощают нерастворимые вещества (макромолекулы, бактерии). Формирующийся пузырек называется фагосомой.
• Пиноцитоз («клеточное питье») – клетки поглощают небольшое количество жидкости, которая может содержать разные растворы. Пузырьки в этом случае обычно меньше, чем фагосомы.
• Рецепторно-опосредованный эндоцитоз. Клетка поглощает чужеродное вещество вместе со своим собственным мембранным рецептором. Связывание лиганда с рецептором запускает сборку комплексов лиганд-рецептор в окаймленные ямки (углубление мембраны, цитоплазматическая поверхность которого покрыта клатрином), а затем окаймленные пузырьки, с помощью которых комплексы лиганд-рецептор проникают в клетку. Здесь клатрин покидает пузырек (последний теперь называется эндосома), а лиганд отделяется от рецептора. Далее эндосома делится на две части: рецептор возвращается на мембрану, лиганд двигается к лизосоме.
4. Экзоцитоз – это выведение веществ из клетки в случае секреции или для освобождения от непереваренного материала. Ограниченный мембраной пузырек или секреторная гранула сливается с мембраной клетки и их содержимое выливается наружу.
5. Компартментализация. Мембрана избирательно блокирует перемещение многих водорастворимых веществ. В цитоплазме находится множество ограниченных мембраной структур – органелл с различным содержимым. Это предотвращает размывание метаболитов и защищает чувствительные реакции от вмешательства веществ извне.
6. Пространственно-временная организация метаболических процессов. Некоторые клеточные мембраны (например, внутренняя мембрана митохондрий или комплекс Гольджи) содержат серии ферментов, организованных так, что в многоступенчатых метаболических процессах промежуточные метаболиты переходят от одного фермента к другому в определенном порядке. Такая организация обеспечивает правильную смену этапов для каждого процесса и задает ограничения скорости процессов, поддерживая постоянную концентрацию метаболитов в том или ином участке клетки.
7. Хранение, транспорт и секреция. Мембранные пузырьки с различным содержимым могут сохраняться в клетке для того, что бы использовать их, когда возникнет необходимость (хранение), могут перемещаться по клетке от одного компартмента к другому для включения в метаболические процессы (транспорт) или выбрасываться из клетки (секреция).
Текучесть мембраны
Мембрана отпочковывается в виде везикулы от органеллы и сливается с другой мембраной, поэтому количество мембраны в органеллах меняется без участия в этом процессов синтеза и разрушения мембраны.
ЦИТОПЛАЗМА
Структуры цитоплазмы делят на 3 труппы:
• Органеллы. Ограничены мембраной, содержат ферменты, каждый вид имеет особое строение и уникальные функции.
• Цитоплазматические включения. Могут быть ограничены мембраной (но не обязательно). По сравнению с органеллами менее постоянны и меньше участвуют в метаболизме.
• Цитоскелет. Это белковая сеть в цитоплазме. Компоненты цитоскелета также формируют отдельные цитоплазматические структуры, например центриоли (из микротрубочек цитоскелета).
Митохондрии
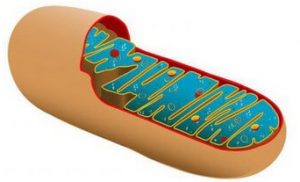
Самые крупные органеллы. Обеспечивают клетку энергией. Обнаружены почти во всех эукариотических клетках. Обычно они равномерно распределены по всей клетке. Однако в клетках, требующих больших затрат энергии, они могут аккумулироваться. Примеры таких клеток – кардиомиоциты, клетки почечных канальцев, в которых происходит активный транспорт ионов. Структура митохондрий
Размер митохондрий сравним с размерами бактерии (2-6 мкм в длину, 0,2 мкм в диаметре), они могут иметь форму сферы, яйца, волокон. Каждая митохондрия ограничена двумя мембранами.
• Наружная мембрана имеет гладкий контур и образует непрерывное, относительно проницаемое покрытие, которое пропускает многие мелкие молекулы.
• Внутренняя мембрана имеет меньше пор и менее проницаема. Она образует складки (кристы), которые направлены внутрь митохондрии. Обычно они имеют форму книжных полок, но у клеток, синтезирующих стероиды, они похожи на трубочки. Внутренняя поверхность крист покрыта внутренними мембранными субъединицами, или F1 субъединицами. Это места активности митохондриальной АТФазы. На внутренней поверхности крист также расположены рибосомы. Во внутренней мембране находятся компоненты электронно-транспортной системы (ферменты, кофакторы), играющие важную роль в работе митохондрий – цитохромы, дегидрогеназы, флавопротеины.
• Между двумя мембранами (наружной и внутренней) образуется межмебранное пространство, которое продолжается в пространство внутри крист. Внутренняя мембрана ограничивает митохондриальный матрикс.
• Митохондриальный матрикс содержит воду, растворенные вещества и матриксные гранулы (содержат ионы кальция), кольцевую ДНК и митохондриальные рибосомы, напоминающие бактериальные. Также в матриксе содержатся многочисленные ферменты, участвующие в цикле Кребса, окислении липидов и синтезе митохондриальной ДНК.
Функции. Митохондрии обеспечивают энергию для химических и механических процессов в клетке. Энергия, образующаяся при окислении органических соединений, запасается в митохондриях в виде молекул АТФ, которые синтезируются за счёт движения электрона по электронно-транспортной цепи белков внутренней мембраны. Затем АТФ покидает митохондрию и запасенная энергия освобождается в различных частях клетки. В митохондрии происходит синтез ДНК и некоторых белков.
РИБОСОМЫ
Это органеллы, синтезирующие белки. Митохондриальные (прокариотические) рибосомы мельче (20 нм), чем цитоплазматические рибосомы эукариот (25 нм).
Структура рибосом
Каждый тип рибосом имеет 2 субъединицы, большую и малую (по константе седиментации при ультрацентрифугировании), у митохондриальной 50S и 30S, у цитоплазматической – 60S и 40S соответственно. Рибосома представляет собой рибосомальную РНК (рРНК), синтезированную в ядрышке клетки, и ассоциированные с ней белки, синтезированные в цитоплазме. Эти компоненты базофильны, при световой микроскопии они выглядят как базофильные частицы (раньше их называли эргастоплазма в железистых клетках и тельца Ниссля в нейронах). В электронном микроскопе рибосомы выглядят как мелкие электронноплотные гранулы в цитоплазме.
Локализация и функции
В цитоплазме рибосомы существуют в двух видах:
• Свободные рибосомы распределены по одной в цитоплазме.
• Полирибосомы (полисомы) – группы рибосом вдоль матричной РНК (мРНК), что позволяет синтезировать множество копий белка с одной мРНК. Рибосомы считывают (транслируют) информацию с мРНК и, таким образом, играют критическую роль в синтезе белка. Полисомы могут находиться в цитоплазме свободно или быть связанными с мембраной гранулярной эндоплазматической сети (ГрЭС). Свободные полисомы синтезируют структурные белки и ферменты для внутриклеточного использования. Полисомы, связанные с ГрЭС, синтезируют белки для секреции (выведения за пределы клетки).
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС)
Это комплекс органелл, участвующих в синтезе, упаковке и обработке различных веществ.
Это свободно анастамозирующая сеть мембран, которая формирует пузырьки или цистерны. Они могут быть удлиненные, плоские, круглые, трубчатые.
Транспортные пузырьки (везикулы) – это маленькие ограниченные мембраной пузырьки, которые отпочковываются от ЭПС и направляются к комплексу Гольджи для дальнейшей обработки и упаковки их содержимого. В зрелых клетках ЭС бывает двух видов, гранулярная (ГрЭПС) и гладкая (ГЭПС).
Структура ГрЭПС
Сеть называется гранулярной из-за расположения на мембранах рибосом. Располагается в цитоплазме и в некотррых местах без перерыва продолжается в наружную мембрану ядерной оболочки. Цистерны ГрЭПС обычно параллельные, плоские и вытянутые, особенно в клетках, специализирующихся на синтезе белка (ацинарные клетки поджелудочной железы, плазматические клетки и т.д.), в которых ГрЭПС очень хорошо развита. В эпителиальных клетках, секретирующих белки, ГрЭПС обычно располагается между базальной плазматичесой мембраной и ядром. Рибосомы придают ГрЭПС базофильные свойства. Структура ГрЭПС хорошо видна только в электронном микроскопе. Уникальные для ГрЭПС белки – docking protein, работающий как рецептор, и рибофорины I и II. Функция
Основная функция ГрЭПС – синтез белков для выведения из цитоплазмы:
• Секреторные белки (коллаген)
• Белки для клеточной мембраны
• Лизосомальные белки
Структура ГЭПС
Сеть не имеет рибосом и выглядит гладкой в электронном микроскопе. Ее цистерны боле трубчатые или везикулярные, чем у ГрЭПС. Окрашивается плохо, в световом микроскопе неотличима от остальной цитоплазмы. Развитая ГЭПС обнаруживается в клетках, синтезирующих стероидные гормоны (кора надпочечников, гонады) и гепатоцитах (метаболизм гликогена, детоксикация токсических веществ). Специфическая ГЭПС – саркоплазматический ретикулум – характерна для скелетных мышц, где она участвует в мышечном сокращении, запасая и выделяя ионы кальция.
Функции
Белок не синтезирует. Содержит много ферментов для метаболизма липидов, синтеза стероидных гормонов, распада гликогена и детоксикации (путем конъюгации, окисления и метилирования токсических продуктов).
КОМПЛЕКС ГОЛЬДЖИ
Участвует в процессах, связанных с секрецией веществ, текучестью мембраны и везикулярном транспорте.
1. Структура
Состоит из:
• Стопки 3-10 отдельных, слегка выгнутых плоских цистерн
• Многочисленных везикул на периферии стопки. Нескольких больших конденсирующих вакуолей у вогнутой поверхности стопки. Выпуклая формирующаяся cis-поверхность обычно находится рядом с ближайшим расширением цистерн ЭПС и окружена транспортными везикулами. Вогнутая созревающая trans-поверхность часто связывает несколько конденсирующих вакуолей и направлена от ядра.
2. Функции
a. Синтез олигосахиридов/полисахаридов с помощью гликозилтрансфераз (инициация, удлиннение/укорочение цепей).
b. Модификация секретируемых продуктов – гликозилирование белков и липидов, сульфатирование гликозаминогликанов (ГАГ), т.е. комплекс Гольджи необходим для синтеза и секреции гликопротеинов, протеогликанов, гликолипидов и сульфатированных ГАГ.
c. Упаковка секретируемых продуктов в везикулы, секреторные везикулы (или секреторные гранулы), которые транспортируются к клеточной мембране для экзоцитоза.
d. Концентрирование и сохранение секретируемых продуктов. В некоторых клетках комплекс Гольджи концентрирует и сохраняет секретируемые продукты перед их секрецией. Эта функция выполняется конденсирующими вакуолями на trans-поверхности комплекса Гольджи. Они также часто служат предшественниками секреторных гранул.
3. Локализация. Комплекс Гольджи обычно расположен рядом с ядром и часто обнаруживается около центриолей (которые также могут играть важную роль в направлении транспорта везикул). Лучше всего комплекс Гольджи развит в нервных и железистых клетках, т.е. клетках, которые специализируются на секреции.
4. Прохождение веществ через комплекс Гольджи. Считается, что секретируемые материалы перемещаются через комплекс Гольджи в одном направлении. Сначала транспортные пузырьки отпочковываются от эндоплазматической сети и сливаются с формирующей cis-поверхностью комплекса Гольджи. Затем содержимое пузырьков модифицируется, по мере того, как оно перемещается от цистерны к цистерне по направлению к созревающей trans-поверхности. От последней затем отпочковываются секреторные пузырьки, содержащие конечный продукт. Однако эта теория сейчас пересматривается. Получены доказательства, что везикулы, ассоциированные с комплексом Гольджи, отличаются по их источнику, точке назначения, функции, содержимому и составу поверхности. Некоторые неклатриновые покрывающие везикулы белки (например, бета-COP) ассоциированы со специфическими областями комплекса Гольджи, что позволяет предположить, что разные типы везикул могут сливаться и отпочковываться и от cis-, и от transповерхностей, и от промежуточных мембран комплекса Гольджи.
ФАГОСОМЫ
Это мембранно-ограниченные пузырьки различных размеров, содержащие материалы для переваривания лизосомами. Известны два основных типа фагосом: гетерофагосомы содержат продукт гетерофагии, то есть материал, имеющий внеклеточное происхождение и поглощенный в результате фагоцитоза. Аутофагосомы – содержат продукт аутофагии, то есть материал внутриклеточный, например, изношенные или поврежденные органеллы. Переваривание содержимого фагосом начинается тогда, когда с ней сольется одна или несколько первичных лизосом, в результате чего образуется вторичная лизосома (описано ниже). Обратите внимание, что некоторые авторы используют термин гетерофагосома для обозначения вторичной лизосомы.
ЛИЗОСОМЫ
Это сферические мембранно-ограниченные пузырьки, которые могут содержать более 50 различных ферментов, действующих как клеточная система пищеварения. Активность этих ферментов отличает лизосомы от других клеточных гранул. Фермент, который наиболее широко используется для идентификации лизосом – кислая фосфатаза, поскольку она встречается практически исключительно в лизосомах. Другие ферменты, которые часто встречаются в лизосомах – рибонуклеазы, дезоксирибонуклеазы, катепсины, сульфатазы, бета-глюкуронидазы, фосфолипазы, а также другие протеазы, липазы и глукозидазы. Врожденный дефицит или повреждение какого-либо фермента лизосом может привести к накоплению субстрата этого фермента в цитоплазме, что может стать опасным для жизни. Обычно ферменты лизосом являются гликопротеинами и наибольшую активность проявляют в кислой среде. Лизосомы имеют различные размеры и электронную плотность в зависимости от их активности.
1. Первичные лизосомы – мелкие (5-8 нм в диаметре), электронно-плотные пузырьки. Они выглядят как черные кружочки на электронных
микрофотографиях. Ферменты таких лизосом неактивны, они служат местом хранения ферментов в клетке. Ферменты лизосом синтезируются на гранулярной ЭПС и транспортируются в комплекс Гольджи для дальнейшего гликозилирования и упаковывания в везикулы. Первичные лизосомы распределены в цитоплазме. Они обнаруживаются в большинстве клеток, но больше всего их в клетках, обладающих фагоцитирующей активностью (макрофаги, нейтрофилы).
2. Вторичные лизосомы – крупнее и имеют меньшую электронную плотность, имеют пестрый вид на электронной микрофотографии. Вторичные лизосомы формируются слиянием одной или нескольких первичных лизосом с фагосомой. Их основная функция состоит в переваривании продуктов гетеро- и аутофагии. Когда ферменты лизосом сливаются с содержимым фагосом, они активируются. В результате переваривания образуются метаболиты для поддержания жизни клетки, ее роста (мелкие молекулы диффундируют в окружающую цитоплазму), обновления органелл. Лизосомальные ферменты также катаболизируют некоторые продукты клеточного синтеза, регулируя, таким образом, количество и качество секретируемого материала. Вторичные лизосомы обнаруживаются во многих клетках, их количество отражает лизосомальную и фагоцитарную активность клетки.
3. Резидуальные тельца – это мембранно-ограниченные включения различных размеров и электронной плотности, ассоциированные с терминальной фазой функционирования лизосом. Они содержат неперевариваемый материал, такой, как пигменты, кристаллы, некоторые липиды. Некоторые клетки (например, макрофаги) выбрасывают резидуальные тельца как мусор, но длительно живущие клетки (нервные, мышечные) накапливают такие тельца. В этом случае содержащие мусор резидуальные тельца отражают возраст клетки, и могут рассматриваться как пигмент «износа» клетки, или гранулы липофусцина. Эти гранулы имеют желтовато-коричневый вид под световым микроскопом и высокую плотность под электронным микроскопом.
ПЕРОКСИСОМЫ
Это мембранно-ограниченные везикулы, содержащие ферменты. Они обычно крупнее первичных лизосом. У некоторых животных (не у человека) они отличаются от лизосом по электронной плотности, гранулярному нуклеоиду уратной оксидазы. Функция пероксисом заключается в метаболизме перекиси водорода. Они содержат уратную оксидазу, Dаминокислотную оксидазу, которые продуцируют перекись водорода, способную убивать бактерии. Они также содержат каталазу, оксидазы которой окисляют различные субстраты и используют удаленный водород в процессе конвертации токсичной перекиси водорода в воду. Пероксисомы также участвуют в глюконеогенезе, помогая бета-окислению жирных кислот. Они распределены в цитоплазме или ассоциированы с гладкой ЭПС.
ДРУГИЕ ЦИТОПЛАЗМАТИЧЕСКИЕ ВКЛЮЧЕНИЯ
Сферические липидные (жировые) капли – видимые включения, служат местами хранения жира (депо), выглядят по-разному в зависимости от способа приготовления гистологического препарата. Гранулы гликогена – также включения, которые PASпозитивны при световой микроскопии и образуют розетки электронно-плотных частиц под электронным микроскопом. И липидные капли, и гранулы гликогена утрачивают окружающую мембрану. Меланин – коричневый пигмент, широко распространенный у позвоночных, часто обнаруживается в электронно-плотных мембранно-ограниченных гранулах, которые называют меланосомами. Меланосом особенно много в клетках эпидермиса кожи и пигментном слое сетчатки.
ЦИТОСКЕЛЕТ
Цитоскелет – это сеть нитевидных элементов, которые могут быть трех типов: микротрубочки (макрофиламенты), микрофиламенты и промежуточные филаменты. Они обеспечивают стабильность структуры для поддержания формы клетки. Это имеет важное значение для движений клетки и реорганизации компонентов ее цитоплазмы. Согласно одной из моделей организации цитоплазмы, органеллы и цитоплазматические включения погружены в нежную сеть, которая называется микротрабекулярная решетка и которая содержит цитоскелет, ферменты и другие компоненты цитоплазмы, которые ранее считались растворенными и случайным образом распределенными в аморфном цитозоле. Эта гипотетическая решетка может обеспечить основу для упорядочивания ферментов, вовлеченных в многоступенчатые реакции (каскады), как это делают некоторые мембраны. Не исключено, однако, что подобная решетка является артефактом, возникающим в процессе фиксации.
1. Микротрубочки
a. Строение. Микротрубочки – это самые толстые компоненты цитоскелета, их диаметр составляет 24 нм. Они представляют собой тонкие трубочки различной длины с плотной стенкой (5 нм толщиной) внутренним просветом диаметром 14 нм. Стенка трубки состоит из субъединиц, которые называются гетеродимерами тубулина. Каждый из гетеродимеров состоит из белков α-тубулина и βтубулина. Гетеродимеры тубулина организованы в нитевидные полимеры, которые называются протофиламентами. Тринадцать таких нитей, уложенных параллельно друг другу, и образуют стенку микротрубочки. Длина микротрубочки увеличивается путем добавления с одной стороны (место нуклеации, или плюс-конец) новых гетеродимеров. Этот процесс можно контролировать экспериментально, регулируя концентрацию ионов кальция или воздействуя антимитотическими алкалоидами. Так, колхицин блокирует процесс связывания новых гетеродимеров в плюс-концом. Винбластин разрушает микротрубочки, связываясь со свободным тубулином.
b. Функция. Микротрубочки играют роль в поддержании формы клетки, аксоплазматическом транспорте в нейронах, распределении меланина в пигментных клетках, движении хромосом во время митоза, организации комплекса Гольджи и челночных перемещениях везикул внутри клетки. В отличие от микрофиламентов, микротрубочки не способны сокращаться. Укорочение микротрубочки происходит за счет отщепления гетеродимеров (деполимеризации) от ее минус-конца.
c. Локализация. Микротрубочки обнаруживаются в цитоплазме большинства клеток и в высоко организованных группах – центриолях, ресничках, жгутиках, базальных тельцах и митотическом веретене деления.
Центриоли
1. Строение. Центриоль – это цилиндрическая группа микротрубочек, имеет средний диаметр 150 нм и среднюю длину 350-500 нм. Состоит из 9 триплетов микротрубочек (9х3), расположенных по косой к стенке трубки. Каждая микротрубочка триплета имеет общую стенку с соседней микротрубочкой этого триплета. В интерфазе клеточного цикла в клетке имеется 2 центриоли, расположенные перпендикулярно друг к другу. Каждая центриоль окружена несколькими электронноплотными спутниками, или перицентриольными тельцами. Другие микротрубочки цитоплазмы расходятся от перицентриольных телец лучами в цитоплазму.
2. Функция. Центриоли – это структурные организаторы клетки. Удвоение центриолей предшествует делению клетки, в процессе митоза центриоли организуют микротрубочки веретена деления. Даже in vitro изолированные центриоли могут контролировать полимеризацию микротрубочек. В клетке центриоли могу переносить неизученные физические организующие силы через микротрубочки радиально от перицентриольных телец. Через воздействие на микротрубочки, центриоли могу контролировать транспорт органелл, везикул и гранул внутри клетки. Центриоли дают начало базальным тельцам (см. ниже).
3. Локализация. Между делениями клетки центриоли расположены ближе к ядру клетки, часто окружены комплексом Гольджи. Центриоли, ассоциированные с комплексом Гольджи, образуют клеточный центр, который виден как светлая область рядом с ядром. В ходе S-фазы интерфазы каждая центриоль удваивается, давая начало процентриоли, которая растет под прямым углом к материнской центриоли. В процессе митоза новая пара центриолей мигрирует к противоположному полюсу клетки для организации веретена деления.
I. Базальные тельца (кинетосомы). В каждой клетке, имеющей реснички или жгутики, центриоли мигрируют к апикальной мембране клетки и дают начало базальным тельцам, примерно так же, как происходит удвоение центриолей при делении клетки. Базальные тельца по строению похожи на центриоли (9 триплетов микротрубочек, 9х3), находятся в цитоплазме, по одному в основании каждой реснички или жгутика, и служат якорем и организатором микротрубочек для этих структур.
II. Ресничка. Клетки с ресничками обычно имеют сотни подвижных ресничек, 5-10 мкм в длину, 0,2 мкм в ширину. Каждая ресничка представляет собой выпячивание клеточной поверхности, покрытое плазматической мембраной. Каждая ресничка имеет сердцевину, или аксонему, состоящую из 9 периферических дуплетов микротрубочек, которые окружают центральную пару одиночных микротрубочек (9х2 + 2). Дуплеты связаны между собой белком нексином (нексиновые мостики), а радиально расположенные спицы связывают дуплеты с центральной капсулой, окружающей две центральные микротрубочки.
Микротрубочки в дуплете называются микротрубочка А (состоит из 13 протофиламентов тубулина) и микротрубочка В (состоит из 11 протофиламентов тубулина, стенка замыкается за счет микротрубочки А). От каждой микротрубочки А к соседнему дуплету отходят 2 динеиновые ручки (динеин – белок с АТФ-азной активностью). Каждая аксонема организована и удерживается базальным тельцем.
III. Жгутик похож на ресничку, но имеет большую длину, и обычно клетка имеет 1-2 жгутика. У человека и других млекопитающих жгутик встречается только у сперматозоидов (хвост), он имеет длину 50-55 мкм и толщину 0,2-0,5 мкм. Аксонема жгутика не отличается по строению от аксонемы реснички.
IV. Веретено деления. Оно образуется между 2 парами центриолей, расположенных на противоположных полюсах делящейся клетки. Некоторые микротрубочки веретена протягиваются от одной центриоли до другой, другие (хромосомные волокна) натянуты между центриолью и центромерой хромосомы. Веретено деления необходимо для разделения хромосом в процессе митоза.
2. Микрофиламенты
a. Строение. Микрофиламенты – самые тонкие компоненты цитоскелета (их толщина 5-7 нм). Обычно они состоят из нескольких типов белка актина. В исчерченных мышечных клетках актиновые филаменты формируют стабильную структуру, связанную с миозиновыми филаментами. Актиновые филаменты в других клетках менее стабильны и могут распадаться (диссоциировать) и повторно собираться. Эти изменения регулируются ионами кальция и циклической АМФ, а также актин-связывающими белками в цитоплазме.
b. Функция. Микрофиламенты обладают сократительной способностью, но для этого они обычно должны взаимодействовать с миозином. В мышечных клетках миозин образует толстые филаменты. В немышечных клетках он существует в растворенном виде. Воздействие цитохолазинами разрушает организацию микрофиламентов и нарушает следующие связанные с ними функции клетки: эндоцитоз, экзоцитоз, сокращение микроворсинок, движения клетки, движения органелл, везикул и гранул, ток цитоплазмы, поддержание формы клетки, разделение дочерних клеток по экватору во время митоза.
c. Локализация. В немышечных клетках микрофиламенты распределены в виде неправильной сети в цитоплазме. Локальные скопления обнаруживаются как тонкая оболочка под плазматической мембраной – терминальная сеть, как параллельные пучки в сердцевине микроворсинок, в цитоплазме у переднего края различных псевдоподий, в ассоциации с другими компонентами цитоплазмы или как ремень (кисет) вокруг экватора делящейся клетки.
3. Промежуточные филаменты
a. Строение. Промежуточные филаменты, как следует из их названия, занимают промежуточное положение по толщине (10-12 нм) между микротрубочками и микрофиламентами. Они состоят из белков, которые структурно близки с ядерными ламинами и, в отличие от микротрубочек и микрофиламентов, которые устроены одинаково во всех типах клеток, промежуточные филаменты имеют разное строение в различных типах клеток. Белки кодируются примерно 70-ю разными генами. Например, цитокератины характерны для эпителиальных клеток, виментин – для клеток мезенхимного происхождения (фибробластов, хондроцитов), десмин – для мышечных клеток, кислый глиальный фибриллярный белок – для клеток глии, нейрофибриллярный белок – для нервных клеток.
b. Функция белков промежуточных филаментов до конца не изучена. Они, вероятно, участвуют в поддержании формы клетки, возможно, как компонент микротрабекулярной сети. Участвуют в образовании межклеточных контактов (десмосом и полудесмосом)*.
Ядерные ламины в настоящее время относятся к V классу промежуточных филаментов, это фибриллярные белки, обеспечивающие структурную функцию и регуляцию транскрипции в ядре клеток.
*Мутации генов кератинов krt5 и krt14 связаны с развитием наследственного кожного заболевания, буллёзного эпидермолиза (Epidermolysis bullosa simplex). При этом заболевании нарушено прикрепление эпидермиса к базальной пластинке, на коже образуются заполненные серозным содержимым пузыри.
c. Локализация. Большая часть промежуточных филаментов распределены в цитоплазме, вокруг ядра клетки они образуют «корзинку», и от нее распространяются на периферию клетки.
ЯДРО
1. Общие сведения. Внешние признаки ядер изменяются от ткани к ткани и от клетки к клетке, но каждое ядро имеет ядерную оболочку, хроматин, нуклеоплазму и одно или несколько ядрышек. Хотя некоторые зрелые клетки теряют ядро (например, эритроциты), как минимум одно ядро присутствует на какой-то стадии развития во всех эукариотических клетках. Ядро содержит линейный код (ДНК) для синтеза компонентов клетки и ее продуктов, наделяющих клетку способностью адаптироваться к изменяющимся условиям окружающей среды и внешним сигналам, таким, как гормоны. Микроскопические признаки ядра важны для идентификации и классификации как здоровых, так и поврежденных клеток и тканей. Ядра демонстрируют высокое разнообразие по: 1) размерам, как абсолютным, так и относительным по сравнению с количеством цитоплазмы (ядерно-цитоплазматическое соотношение), 2) количеству на клетку, позволяющему классифицировать клетку как безъядерную, одноядерную, двуядерную или многоядерную, 3) количеству и распределению хроматина и 4) локализации (базальной, центральной, эксцентричной).
2. Ядерная оболочка. Ядерное содержимое отделено от остальной цитоплазмы двойной мембраной, которая называется ядерной оболочкой. Щель между двумя мембранами узкая (40-70 нм), она называется перинуклеарная цистерна, или перинуклеарное пространство. Ядерная оболочка часто рассматривается как выпячивание гранулярной ЭПС, поскольку ее наружная поверхность усыпана рибосомами и имеет связи с гранулярной ЭПС. Внутренняя поверхность внутренней мембраны покрыта фиброзной пластинкой – слоем, состоящим из ядерных белков промежуточных филаментов – ламинов. Ядерная оболочка перфорирована многочисленными отверстиями – ядерными порами, каждая из которых имеет диаметр около 70 нм и окружена 8 глобулярными субединицами, аннулярными белками, которые имеют вид восьмиугольника на некоторых препаратах. Каждая пора покрыта тонкой белковой диафрагмой. Поры образуют каналы для движения важных молекул между ядром и цитоплазмой, в том числе нуклеиновых кислот, синтезируемых в ядре, а используемых в цитоплазме (мРНК, рРНК, тРНК), и белков, синтезируемых в цитоплазме, а используемых в ядре (гистоны, полимеразы).
3. Хроматин – интенсивно базофильный, состоит из ДНК и ассоциированных с ней гистонов и негистоновых белков.
- Нуклеосомы. Изолированный хроматин виден на электронных микрофотографиях как тонкие нити с выступающими частицами в виде шариков через регулярные промежутки. Каждая нить – это двойная спираль молекулы ДНК, а частицы – повторяющиеся структурные субъединицы хроматина, которые называются нуклеосомами. Каждая нуклеосома состоит из 166 пар оснований нити ДНК, скрученной вокруг сердцевины из 8 гистонов (по 2 копии каждого: Н2А, Н2В, Н3, Н4). Часть нити ДНК между двумя нуклеосомами содержит дополнительно 48 пар оснований и называется линкерным регионом (линкерной ДНК, линкером). Другой гистон (обычно Н1) связан с наружной поверхностью нуклеосомы и линкера. Цепочка из нуклеосом сворачивается в супер-спираль (6 нуклеосом на виток), так образуется конденсированный хроматин – гетерохроматин.
- Типы хроматина. Ядро содержит высоко конденсированный хроматин (гетерохроматин), который окрашивается в темный цвет основными красителями. Поскольку для транскрипции с ДНК она должна быть раскручена клетки с темно окрашенными ядрами (содержащими гетерохроматин) менее активны в транскрипции ДНК и используют меньшую часть их общего генома по сравнению с другими клетками. Несвернутый хроматин (эухроматин) окрашивается плохо и его трудно различить даже в электронный микроскоп. Большие, бледно окрашенные ядра (эухроматин) обычно бывают у клеток с высокой активностью транскрипции и быстрыми клеточными делениями.
- Паттерны хроматина. Количество и распределение ядерного хроматина часто импользуют для идентификации клеточного типа, особенно для клеток, которые не имеют характерного окрашивания цитоплазмы. Даже в большинстве эухроматических ядер ободок гетерохроматина часто обнаруживается на внутренней поверхности ядерной оболочки в ассоциации с фиброзной пластинкой. Этот связанный с ядерной оболочкой гетерохроматин позволяет увидеть границы ядра в световой микроскоп.
- Хромосомы, наиболее конденсированная форма хроматина, становятся видны во время митоза. Супер-спирали гетерохроматина образуют складки и накручиваются на центральную основу из негистоновых белков. В клетках человека имеется 46 хромосом, 44 соматических хромосомы образуют 22 пары структурно похожих хромосом. Одна пара половых хромосом состоит из двух различных (XY) хромосом у мужчин и двух похожих (XX) хромосом у женщин. У женщин используется только одна из пары Х-хромосом. Неактивная Х-хромосома часто видна как глыбка гетерохроматина, которую называют половой хроматин, или тельце Барра. В большинстве клеток тельце Барра находится у внутренней поверхности ядерной оболочки. В нейрофилах оно может появиться как отросток от сегментированного ядра в виде барабанной палочки.
- Кариотип. Кариотип клетки – это перечень ее хромосом или изображение ее хромосом, разложенных по типам. Приготовление такого изображения называется кариотипированием. Клетки в культуре стимулируют к митозу фитогемагглютинином (митоген растительного происхождения). Делящиеся клетки обрабатывают колхицином (вспомните механизм и точку приложения действия колхицина), чтобы остановить митоз в метафазе, когда хромосомы высоко конденсированы и хорошо видимы. Лизис клеток с помощью гипотонического раствора позволяет хромосомам распределиться на стекле с минимальным наложением друг на друга или без такого наложения. Далее хромосомы фотографируют, собирают в пары и располагают в определенной последовательности. Кариотипирование позволяет каталогизировать хромосомы для выявления их структурных нарушений, потери или появления избыточных хромосом.
4. Ядрышко. В ходе интерфазы (между митозами) каждое ядро обычно имеет как минимум одно интенсивно базофильное тело, которое называется ядрышком. Ядрышки – места синтеза большей части рРНК. Обычно они отличаются от гетерохроматина, но в темных ядрах могут быть видны плохо. Наиболее многочисленные и крупные ядрышки встречаются в эмбриональных клетках, в клетках, активно синтезирующих белки, и в быстро растущих злокачественных опухолевых клетках. Небольшие количества гетерохроматина прикрепляются к ядрышкам. Значение такого ядрышкоассоциированного хроматина неизвестно. При подготовке клетки к митозу ядрышки исчезают и снова появляются после завершения митоза. Под электронным микроскопом можно увидеть различные компоненты ядрышка.
a. Аморфная часть – бледно окрашенная часть ядрышка, содержащая ядрышковый организатор ДНК, который несет код для рРНК. У человека 5 пар хромосом имеют имеют регионы ядрышкового организатора. Таким образом, возможно образование 10 ядрышек, но слияние ядрышковых организаторов приводит к появлению меньшего числа более крупных ядрышек. Новая синтезированная рРНК появляется первой в этом регионе.
b.Нуклеолонема – термин, который используют для обозначения нитевидной базофильной структуры ядрышка. Нуклеонема содержит 2 компонента, богатых рРНК и различимых в ЭМ.
- Фиброзная часть состоит из плотно упакованных рибонуклеобелковых волокон, 5-10 нм в диаметре. Эти волокна состоят из вновь синтезированных первичных транскриптов генов рРНК и ассоциированных с ними белков. Вновь синтезированная рРНК появляется второй в этом регионе.
- Гранулярная часть, 15-20 нм в диаметре, которые представляют собой созревающие рибосомальные субъединицы в процессе сборки для экспорта в цитоплазму. Вновь синтезированная рРНК появляется третьей в этом регионе.
5. Нуклеоплазма. Это матрикс, в котором находятся другие внутриядерные компоненты. Он состоит из ферментных и неферментных белков, метаболитов, ионов и воды. Он включает ядерный матрикс – фибриллярную ядерно-скелетную структуру, которая появляется, чтобы связывать некоторые рецепторы гормонов и вновь синтезированную ДНК
ФУНКЦИИ КЛЕТКИ
Способность клеток выполнять свои функции зависит от доступных им ресурсов. Количество и типы энергии, продукты обмена и их удаление, информация, закодированная в генах, внутренние и внешние факторы, которые контролируют их доступ к этой информации, – основные факторы, которые определяют функции клетки. Растущие клетки и клетки при регенерации используют большую часть ресурсов клетки на подготовку к митозу и последующую дифференцировку и рост. Полностью дифференцированные клетки обычно концентрируются на выполнении более специализированных функций, таких как секреция или сокращение. Поддержание постоянства внутренней среды (гомеостаза) даже в явно покоящихся клетках, требует расходов значительного количества энергии и других ресурсов.
1. Репродукция клеток. Цикл воспроизведения клеток называется клеточным циклом. Каждый полный клеточный цикл заканчивается делением клетки (митозом), в результате которого образуется 2 дочерние клетки, которые обычно имеют размеры в два раза меньше, чем материнская клетка.
a.Митоз и интерфаза. Ранние сведения о клеточной репродукции фокусировались на легко выявляемых структурных изменениях в ходе митоза. По всей видимости, неактивная фаза между последующими митозами казалась тогда периодом покоя и поэтому была названа интерфазой. Даже в быстро делящихся клетках время митоза намного короче времени, которое клетка проводит в интерфазе. Сейчас мы знаем, что в интерфазе происходит множество важных событий, включая восстановление после митоза и подготовку к следующему митозу. В настоящее время и митоз, и интерфаза рассматриваются как сложные и важные компоненты клеточного цикла, и каждый подразделяется на несколько этапов для облегчения понимания событий клеточной репродукции.
b.Этапы клеточного деления (митоза). Митоз – это короткий непрерывный процесс, который делят на 4 фазы: профаза, метафаза, анафаза, телофаза.
c. В профазу хроматин интенсивно конденсируется, образуются хромосомы. Ядрышковый организатор наматывается в соответствующую хромосому, ядрышки диспергируют и начинают исчезать. Ядерная мембрана остается интактной. 2 пары центриолей мигрируют к противоположным полюсам клетки, между ними начинается сборка веретена деления.
d.В ходе метафазы ядрышки и ядерная оболочка исчезает. Хромосомы выстраиваются вдоль экватора клетки между двумя парами центриолей, каждая хромосма расщепляется вдоль для формирования пары сестринских хроматид. Каждая хромосома имеет центромеру (кинетохор), к которой прикрепляются микротрубочки веретена деления.
e.В ходе анафазы сестринские хроматиды отделяются друг от друга и двигаются к противоположным полюсам вдоль веретена деления. За счет того, что первой двигается центромера, хроматида приобретает форму буквы «V».
f. В процессе телофазы хромосомы начинают раскручиваться. Ядрышки и ядерные оболочки появляются как компоненты двух отдельных ядер на противоположных полюсах клетки. Пучки микрофиламентов, расположенные под мембраной клетки, образуют сужение в области экватора клетки. Затягивание этого сужения постепенно делит цитоплазму и органеллы между дочерними клетками.
2. Клеточный цикл. Модель клеточного цикла принимает во внимание важные, но менее видимые изменения, происходящие в клетке между клеточными делениями. Он включает 4 фазы митоза, но фокусируется на времени синтеза ДНК и подразделяется на 3 фазы: G1, S и G2.
a. Фаза G1 (gap 1) клеточного цикла следует за телофазой митоза. Это период, в котором не происходит синтез ДНК, но синтезируются РНК и белки, и каждая дочерняя клетка достигает размеров материнской клетки. Обычно это самая продолжительная фаза клеточного цикла, и самая отличающаяся по времени в зависимости от типа клетки. В быстро делящихся клетках, например, эмбриональных или опухолевых, эта фаза короткая и быстро переходит в следующую. Более высоко дифференцированные клетки могут выйти из клеточного цикла и вступить в фазу G0, в которой подготовка к митозу приостановлена, а клетка выполняет специализированные функции. Некоторые клетки, вступившие в G0, больше неспособны вернуться в клеточный цикл (мышечные, нервные), это – терминально дифференцированные клетки. Другие клетки в G0 (гепатоциты, фибробласты) могут вновь вернуться в клеточный цикл в процессе регенерации в ответ на повреждение.
b.В ходе S фазы происходит репликация ДНК и удвоение центриолей.
c.В ходе G2 фазы (gap 2) происходят окончательные приготовления к делению клетки – синтез тубулина для клеточного веретена, накопление АТФ для энергозатратного митоза. Во время митоза синтетические процессы проходят на минимальном уровне.
3. Дифференцировка клеток. Совершенствование структуры и функции клеток происходят во время эмбрионального и фетального развития, а также после рождения. Процесс дифференцировки клеток приводит в целом к менее частым их делениям и выполнению меньшего количества функций, но более высокого качества. О функции клеток можно приблизительно судить по содержащимся в них органеллам. Так, клетки, которые специализируются на синтезе белка, имеют хорошо развитые гранулярную ЭПС и комплекс Гольджи. Дифференцировка клеток может привести к драматическим изменениям, поэтому она не происходит внезапно. Она совершается через серию шагов, часто отделенных один от другого одним или более прохождением через клеточный цикл, и включает взаимодействия между клеточным микроокружением, метаболическими сдвигами в цитоплазме и информацией в ее ДНК.
4. Межклеточные взаимодействия. Ткани, органы и системы органов – это собрания клеток и их продуктов, которые могут действовать вместе, выполняя сложные функции. Взаимодействия между клетками разделяют на прямые и непрямые.
a. Прямые взаимодействия. В некоторых тканях, особенно эпителиальных, клетки имеют прямые контакты друг с другом довольно на большом протяжении их клеточной мембраны. Эти области контактов часто отмечены специальными структурами плазматической мембраны, которые называют соединительными комплексами. Некоторые компоненты таких комплексов специализируются на прикреплении (физическое взаимодействие), тогда как другие (например, щелевые контакты), обеспечивают передачу электрических или химических сигналов от клетки к клетке через специальные цитоплазматические каналы.
b. Непрямые взаимодействия. Сигналы от одной клетке к другой могут передаваться и в том случае, если между клетками нет физического контакта. Такие сигналы могут осуществляться на расстоянии с помощью гормонов или других сигнальных молекул. Иногда (распознавание клеток друг другом, контактное подавление клеточного деления) передача такого сигнала требует временного физического контакта.