ОБЩАЯ ХАРАКТЕРИСТИКА ЭНДОКРИННОЙ СИСТЕМЫ
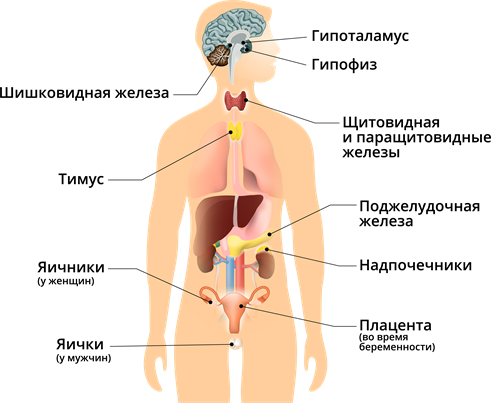
Компоненты. Эндокринная система включает в себя органы (гипофиз, щитовидная железа, надпочечники), скопления эндокринных клеток в экзокринных железах (островки Лангерганса), а также изолированные эндокринные клетки (клетки системы DNES в слизистой оболочке пищеварительного тракта)
Эмбриональные источники развития. Эндокринные железы не имеют протоков и изначально представляют собой эпителиальные впячивания, например, эктодермы ротовой полости или энтодермы кишки. Впоследствии, они обособляются и теряют контакт с «родительским» эпителием, образуя отдельный орган.
Микростроение. Для эндокринных желёз типично наличие секреторных клеток, которые образуют тяжи, скопления или фолликулы и находятся в тесном контакте с большим количеством капилляров или синусоидов.
Виды секрета. Эндокринные железы выделяют свой секрет, обычно гормоны, непосредственно в кровоток. Другие вещества, выделяемые непосредственно в кровоток, такие как ферменты, сывороточные альбумины и т.д., считаются эндокринными секретами. Гормоны – молекулы, обладающие специфическими регуляторными эффектами на клетки-мишени, находящиеся, как правило, в отдалении от железы. Гормоны оказывают выраженные эффекты в очень незначительных концентрациях. Они прямо или опосредованно влияют на все ткани и являются важнейшими регуляторами гомеостаза в организме. Гормоны регулируют метаболизм белков, жиров и углеводов, водный и минеральный баланс, рост, первичные и вторичные половые признаки, поведение, темперамент и эмоции.
Пептидные гормоны являются белками, гликопротеинами или короткими пептидными цепочками, связывающимися со специфическими рецепторами на поверхности клеток-мишеней. Обладают способностью стимулировать продукцию клеткамимишенями внутриклеточных вторых посредников, таких, как цАМФ.
Стероидные гормоны являются жирорастворимыми, легко проникают через мембрану клетки-мишени и связываются со специфическими цитоплазматическими и ядерными протеинами. Ядерные рецепторы последовательно связываются с ДНК и оказывают прямое воздействие на транскрипцию генов.
Нейроэндокринная система. Сложные взаимодействия клеток, тканей и органов контролируются и координируются двумя системами: нервной и эндокринной. Всё чаще эти две системы рассматривают как части единой нейроэндокринной системы. Гипофиз, ранее считавшийся главной железой благодаря его способности контролировать функции других желёз, на сегодняшний день считается местом соединения эндокринной и нервной систем. Секреторная активность обеих его частей, аденогипофиза и нейрогипофиза, находятся под контролем расположенной поблизости части мозга – гипоталамуса. Активность гипоталамуса регулируется нервными связями с другими отделами мозга, а также посредством отрицательной обратной связи с гормонами, продуцируемыми клеткамимишенями гипофиза. Заболевания гипофиза проявляются чаще всего гипосекрецией или гиперсекрецией продуцируемых гормонов и могут быть следствием опухолей самого гипофиза, его органов-мишеней, а также гипоталамуса.
ГИПОФИЗ
Гипофиз располагается в ямке турецкого седла тела клиновидной кости позади зрительного перекрёста и соединяется с гипоталамусом, частью промежуточного мозга, с помощью воронки. Состоит из двух основных частей: передней – аденогипофиза и задней – нейрогипофиза, которые отличаются по происхождению, строению и функциям
Общее строение и эмбриональное происхождение гипофиза
Аденогипофиз
Эмбриональное происхождение Выпячивание эктодермы ротовой бухты, направленное краниально. Контактирует и сливается с нейрогипофизом.
Общее строение Состоит из тяжей железистых эпителиальных клеток, между которыми располагаются синусоиды вторичного капиллярного сплетения. Не имеет прямой иннервации нейронами гипоталамуса, иннервируется автономными нервами сплетения внутренней сонной артерии.
Отделы:
Передняя доля (дистальная часть) – большая часть гипофиза;
Туберальная часть – верхнее выпячивание дистальной части, которое охватывает воронку нейрогипофиза; Промежуточная доля – узкий участок на границе с нейрогипофизом.
Нейрогипофиз
Эмбриональное происхождение Выпячивание нейроэктодермы промежуточного мозга, направленное каудально, считается частью головного мозга
Общее строение Состоит из множества аксонов, тела которых располагаются в супраоптическом и паравентрикулярном ядрах гипоталамуса
Отделы:
Воронка – состоит из срединного возвышения и ствола, проводит нейроны гипоталамуса к pars nervosa, содержит петли первичного капиллярного сплетения; срединное возвышение серого бугра формирует дно гипоталамуса. Задняя доля (рars nervosa) – расширенная часть нейрогипофиза, содержит терминали аксонов и капилляры.
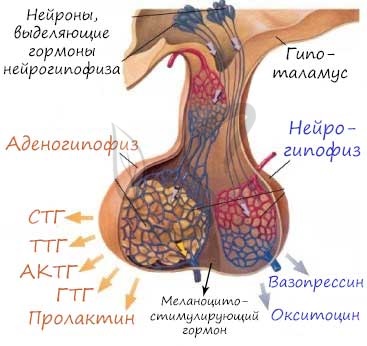
АДЕНОГИПОФИЗ
Секреторные клетки аденогипофиза синтезируют и накапливают один из следующих гормонов: два гонадотропных гормона: фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ), тиреотропный гормон (ТТГ), адренокортикотропный гормон (АКТГ), гормон роста (ГР), или соматотропный гормон (СТГ), и пролактин. Эти гормоны контролируют секреторную активность многих других желёз организма человека. Специфические стимулирующие (либерины) и ингибирующие (статины) гормоны гипоталамуса (релизинг-факторы, или релизинг-гормоны) поступают к клеткам аденогипофиза по системе портальных сосудов и регулируют высвобождение соответствующих гормонов этими клетками.
А. Передняя доля (дистальная часть)
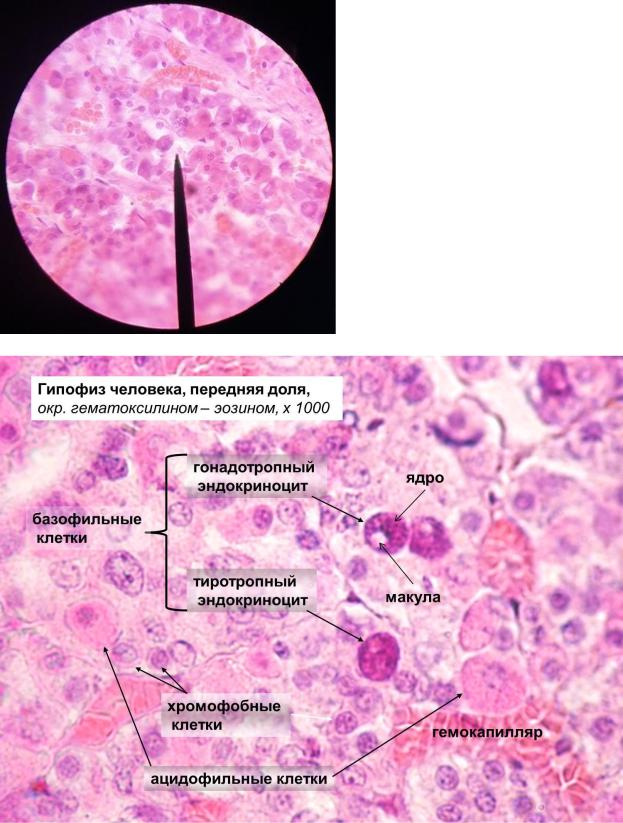
- Хромофобы – слабо окрашивающиеся клетки, на срезах выглядят прозрачными или белесыми. Хромофобы составляют около 50 % эпителиальных клеток передней доли гипофиза. Среди этих клеток выделяют три субпопуляции: 1) – недифференцированные несекреторные клетки, которые могут быть стволовыми; 2) – частично дегранулировавшие хромофилы; 3) – фолликулярные клетки, формирующие поддерживающий каркас для хромофилов, составляют большинство хромофобных клеток, имеют звёздчатую форму, могут обладать способностью к фагоцитозу.
- Хромофилы – секреторные клетки, окрашиваются интенсивно за счёт большого количества цитоплазматических гранул, в которых накапливаются гормоны. Для каждого гормона существуют специфические продуцирующие их клетки. Хромофилы делятся на два класса:
Ацидофильные клетки секретируют простые белки, интенсивно окрашиваются эозином. Располагаются по периферии железы, более мелкие, чем базофильные клетки, однако содержат большее количество более крупных гранул. Ацидофильные клетки включают два основных типа гормонпродуцирующих клеток: соматотропы, продуцирующие гормон роста (соматотропин), и маммотропы продуцирующие пролактин.
Базофильные клетки секретируют гликопротеиды, окрашиваются гематоксилином и другими основными красителями. Располагаются преимущественно в центре, более крупные по сравнению с ацидофильными клетками, но содержат меньше секреторных гранул. Три группы базофильных клеток продуцируют четыре основных гормона. Существуют гонадотропные клетки двух видов: одни продуцируют фолликулостимулирующий гормон (ФСГ), другие – лютеинизирующий гормон (ЛГ). Кортикотропы синтезируют адренокортикотропный гормон (АКТГ). Тиротропные клетки производят тиреотропный гормон (ТТГ).
Б.Туберальная часть
Имеет форму трубки, окружающей воронку нейрогипофиза. Большая часть клеток туберальной части представлена гонадотропами, секретирующими гонадотропные гормоны ФСГ и ЛГ. Клетки располагаются в виде тяжей рядом с кровеносными капиллярами.
В. Промежуточная доля
Развивается из дорсальной части кармана Ратке, у человека является рудиментарной. Состоит из тяжей и фолликулов базофильных клеток, секретирующих меланоцитстимулирующий гормон.
Г. Кровоснабжение гипофиза осуществляется из внутренней сонной артерии.
- Первичное капиллярное сплетение – скопление капилляров, расположенное в области воронки и срединного возвышения, продолжается в туберальную часть. Сплетение получает кровь из передних и задних верхних гипофизарных артерий (Виллизиев круг) и опорожняется в гипофизарные портальные вены.
Гипофизарные портальные вены – мелкие вены и венулы, располагающиеся в средней и нижней части воронки и частично в туберальной части. Получают кровь из первичного капиллярного сплетения и несут её непосредственно во вторичное капиллярное сплетение дистальной части аденогипофиза. Сосуды, несущие кровь напрямую из одного капиллярного сплетения в другое, минуя системный кровоток, называются портальными.
Вторичное капиллярное сплетение – богатая сеть фенестрированных капилляров и синусоидов, расположенная в области передней доли гипофиза, распространяется также в промежуточную долю, имеет сообщения с сосудами задней доли. Синусоиды, находящиеся между фолликулами и тяжами клеток в передней доле, также относятся к этому сплетению и получают венозную кровь непосредственно из гипофизарных портальных вен и артериальную кровь из передних нижних гипофизарных артерий. Кровь из сплетения оттекает по нижним гипофизарным венам в систему внутренних ярёмных вен.
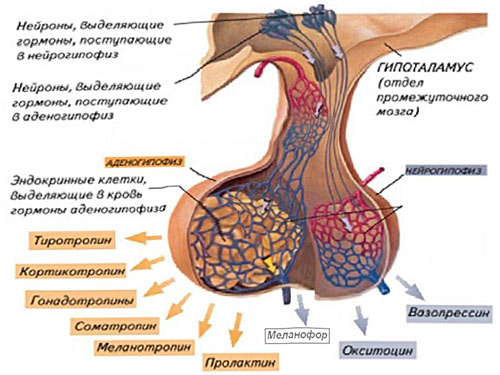
Д. Гормоны гипоталамуса – пептиды низкой молекулярной массы, секретируемые ядрами гипоталамуса, и по аксонам транспортируемые в первичную капиллярную сеть. По портальным венам они поступают во вторичное капиллярное сплетение, а затем к клетками аденогипофиза, стимулируя или подавляя синтез гормонов.
Ядрами в центральной нервной системе называют скопления тел нервных клеток (нейронов), объединенных той или иной функцией. Аксон – отросток нервной клетки, несущий импульс от тела нейрона к клетке-мишени.
Стимулирующие гормоны (либерины, релизинг-гормоны).
- Кортикотропин-релизинг гормон (КРГ), или кортиколиберин – пептид, синтезируемый в паравентрикулярных ядрах гипоталамуса, стимулирует кортикотропные клетки аденогипофиза высвобождать АКТГ.
Гонадотропин-релизинг гормон (ГнРГ), или гонадолиберин – пептид, синтезируемый преоптическим и аркуатным ядрами гипоталамуса, стимулирует высвобождение ФСГ и ЛГ.
Тиротропин-релизинг гормон (ТРГ) или тиролиберин – пептид стимулирующий синтез ТТГ.
Пролактолиберин – гормон, стимулирующий синтез пролактина в аденогипофизе.
Соматолиберин – гормон, стимулирующий синтез СТГ в аденогипофизе.
Ингибирующие гормоны.
Соматостатин, или гормон роста-ингибирующий гормон – подавляет секрецию гормона роста, а также инсулина, глюкагона и других гормонов, ассоциированных с желудочно-кишечным трактом.
Допамин, или пролактин-ингибирующий гормон (ПИК) – нейротрансмиттер, синтезируемый в аркуатном ядре гипоталамуса, подавляет секрецию пролактина.
Секреция других гормонов аденогипофиза подавляется избытком гормонов, секретируемых клетками-мишенями соответствующих эндокринных желез (коры надпочечников, половых желез, щитовидной железы) по механизму отрицательной обратной связи.
Е.Продукция гормонов аденогипофиза
Нейроны ядер гипоталамуса секретируют релизинг-гормоны или ингибирующие гормоны и упаковывают их в нейросекреторные пузырьки.
Аксоны нейронов по гипоталамо-гипофизарному тракту транспортируют эти пузырьки к терминальным расширениям вокруг капилляров первичной капиллярной сети.
Сигналы от нервной системы или уровень гормонов, секретируемых железами-мишенями аденогипофиза по механизму отрицательной обратной связи приводят к высвобождению соответствующих ингибирующих или релизинг-гормонов.
Ингибирующие или релизинг-гормоны поступают в капилляры первичной капиллярной сети, а затем по портальным венам во вторичное капиллярное сплетение.
Здесь они диффундируют через фенестры и стимулируют или подавляют синтез гормонов аденогипофиза соответствующими клетками.
Гормоны аденогипофиза поступают во вторичную капиллярную сеть, покидают аденогипофиз через передние нижние гипофизарные вены и попадают в системный кровоток.
НЕЙРОГИПОФИЗ
Все отделы нейрогипофиза имеют схожее микроскопическое строение. В этой части рассмотрим строение задней доли (pars nervosa), которая состоит из трёх основных компонентов: аксонов, капилляров и питуицитов.
А. Аксоны нейросекреторных клеток. Нейрогипофиз окрашивается слабо, содержит множество немиелинизированных аксонов нейронов, тела которых располагаются преимущественно в супраоптическом и паравентрикулярном ядрах гипоталамуса. Аксоны, идущие от этих ядер к задней доле гипофиза, формируют гипоталамо-гипофизарный тракт. Эти аксоны содержат нейросекреторные пузырьки и имеют терминальные расширения – тельца Герринга, в которых накапливается секрет. Нейросекреторные пузырьки содержат следующие вещества:
Гормоны нейрогипофиза. Нейроны гипоталамуса, оканчивающиеся в нейрогипофизе, высвобождают окситоцин и антидиуретический гормон (АДГ). Окситоцин – пептид, синтезируемый преимущественно паравентрикулярными ядрами гипоталамуса, стимулирует выделение молока молочными железами и сокращение гладких мышц матки во время родов и менструации. АДГ (вазопрессин) – пептид синтезируемый нейронами супраоптических ядер, стимулирует реабсорбцию воды в собирательных трубочках почки и сокращение гладких мышц стенки кровеносных сосудов.
Нейрофизины – связывающие белки, которые образуют комплексы с гормонами нейрогипофиза.
Аденозинтрифосфат (АТФ) – источник энергии для нейросекреции.
Б. Сплетение фенестрированных капилляров, окружающее терминали аксонов нейрогипофиза, доставляет нейросекреторные продукты в системный кровоток.
В. Питуициты – сильно ветвящиеся глиальные клетки, отростки которых окружают и поддерживают немиелинизированные аксоны. Их ядра обычно крупнее и более эухроматичны, чем ядра окружающих фибробластов.
Г. Продукция гормонов нейрогипофиза. Нейроны супраоптического и паравентрикулярных ядер гипоталамуса синтезируют АДГ и окситоцин соответственно. В нейронах эти гормоны упаковываются вместе с нейрофизинами и АТФ в нейросекреторные пузырьки. Нейросекреторные пузырьки транспортируются по аксонам, образующим гипоталамогипофизарный тракт, к терминальным расширениям, располагающимся среди капилляров нейрогипофиза. В ответ на адекватную стимуляцию возникает потенциал действия и происходит экзоцитоз содержимого нейросекреторных пузырьков в терминалях. Высвобождаемые гормоны попадают в капилляры нейрогипофиза и покидают гипофиз через задние нижние гипофизарные вены, попадая в системный кровоток.
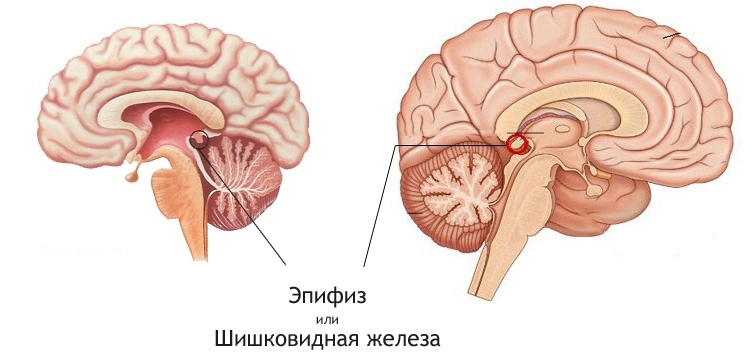
ЭПИФИЗ (ШИШКОВИДНОЕ ТЕЛО)
Небольшой (3-5 мм×5-8 мм) конусовидный орган, соединяется при помощи ножки с крышей промежуточного мозга, в задней части третьего желудочка. Покрыт мягкой мозговой оболочкой с отходящими от неё септами, содержащими кровеносные сосуды и безмиелиновые нервные волокна, которые делят железу на дольки. Эпифиз содержит округлые участки базофильного кальцинированного матрикса, известного как «мозговой песок», размеры и количество которого увеличиваются с возрастом. Рентгеноконтрастность этих отложений и центральное расположение эпифиза в черепе служат хорошим ориентиром для врачей-радиологов. Существует два основных типа клеток эпифиза: пинеалоциты и астроглиальные клетки.
А. Пинеалоциты
Строение. Клетки с большим, неправильной формы, ядром, выраженным ядрышком и бледной базофильной цитоплазмой. При окрашивании серебром видны длинные цитоплазматические отростки, оканчивающиеся расширениями в области септ, рядом с кровеносными капиллярами. Роль иннервации шишковидного тела до конца не ясна.
Функция пинеалоцитов заключается в синтезе мелатонина. Циклические изменения уровня мелатонина в плазме крови в ответ на изменение освещения передаются нейронам, тела которых лежат в супрахиазматических ядрах гипоталамуса. Мелатонин способствует устанавливанию сезонных и циркадных ритмов, оказывает антигонадотропный эффект, задерживая половое созревание до пубертата, обладает противоопухолевой активностью.
Патологические изменения. Опухоли эпифиза встречаются чаще всего у мальчиков и могут приводить к задержке или наоборот раннему половому созреванию. Из-за особенностей расположения опухоли эпифиза могут нарушать циркуляцию ликвора через Сильвиев водопровод, приводя к гидроцефалии.
Б. Астроглиальные клетки, известные также как интерстициальные клетки, имеют удлиненное гетерохроматичное ядро и длинные цитоплазматические отростки, содержащие промежуточные филаменты. Располагаются между кровеносными капиллярами и скоплениями пинеалоцитов.
ОСОБЕННОСТИ СТРОЕНИЯ ЭНДОКРИННЫХ КЛЕТОК
Микроструктура эндокринных клеток напрямую зависит от вида секретируемого гормона. Например, клетки, синтезирующие стероидные гормоны, содержат выраженную гладкую эндоплазматическую сеть, тогда как в цитоплазме клеток, выделяющих пептидные гормоны, более выражена гранулярная эндоплазматическая сеть.
НАДПОЧЕЧНИКИ
Надпочечники располагаются в виде «шапочки» у верхнего полюса каждой почки, состоят из коркового и мозгового вещества, которые имеют разное эмбриональное происхождение, строение и функции.
А. Кора надпочечников
- Эмбриональное происхождение. Кора надпочечников развивается из промежуточной мезодермы.
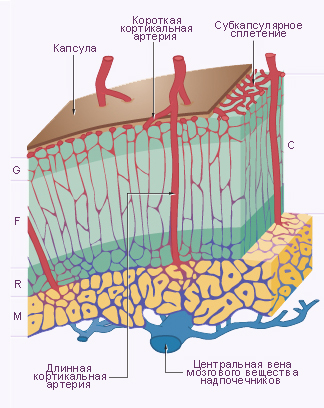
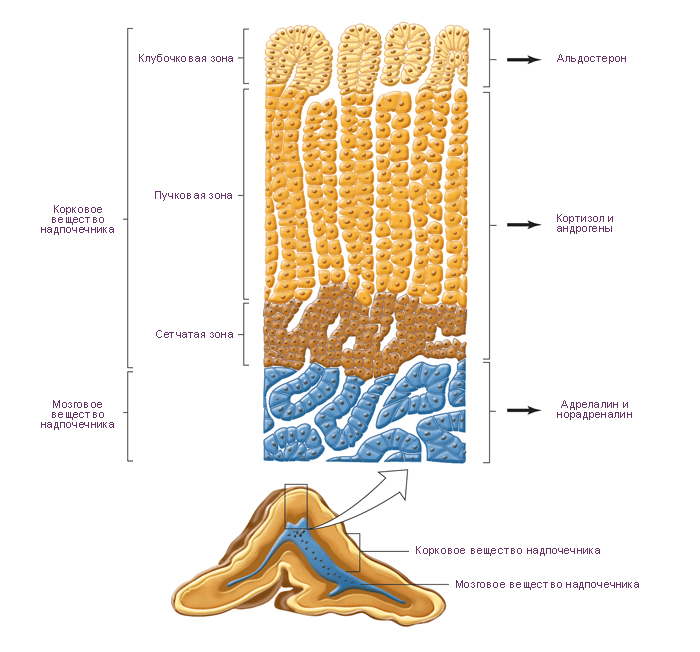
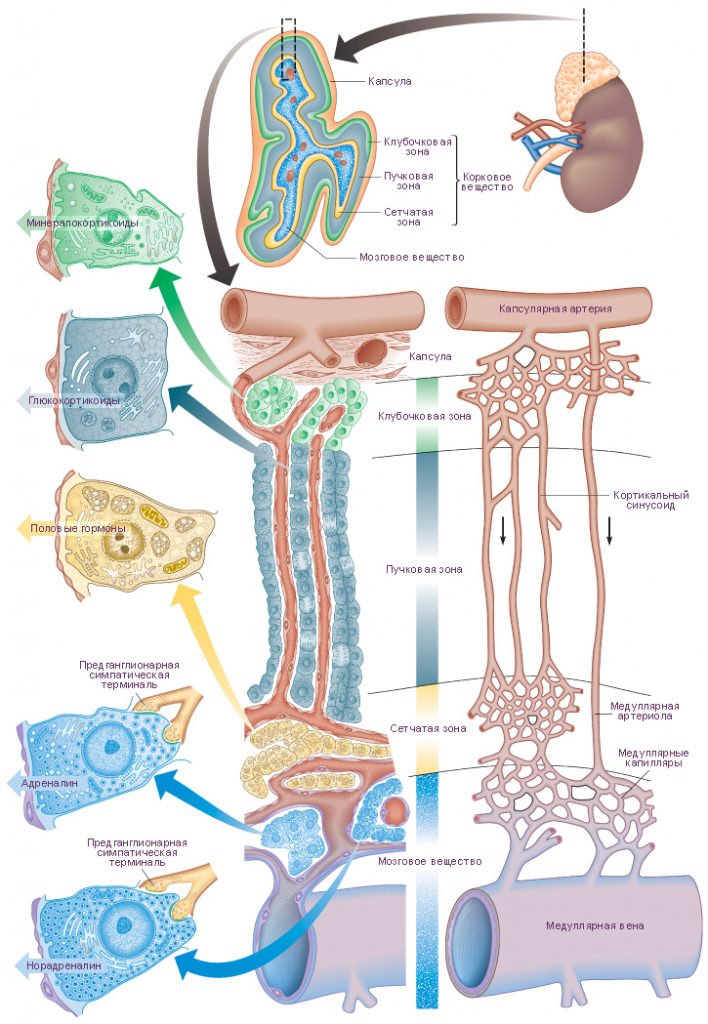
Строение у взрослых. Клетки коры надпочечников имеют строение, типичное для клеток, синтезирующих стероидные гормоны. В коре надпочечников выделяют три зоны: клубочковая зона, пучковая зона и сетчатая зона.
Клубочковая зона располагается по периферии коры, непосредственно под капсулой, и составляет примерно 15 % объема железы. Клетки этой зоны образуют дугообразные скопления (клубочки), окружённые капиллярами, и синтезируют минералокортикоиды.
Пучковая зона – средняя часть коры, составляет примерно 65 % объёма железы. Клетки этой зоны образуют тяжи, расположенные перпендикулярно к поверхности органа, синтезируют глюкокортикоиды и некоторое количество надпочечниковых андрогенов.
Сетчатая зона является самой внутренней частью коры надпочечников и составляет 7 % объёма органа. Клетки располагаются тяжами неправильной форы и образуют сеть. Клетки этой зоны более мелкие и более ацидофильные, чем клетки пучковой зоны, а также содержат меньше жировых капель, больше митохондрий и гранул липофусцина. Сетчатая и пучковая зона могут составлять единую функциональную зону, где большая часть глюкокортикоидов и андрогенов будет синтезироваться клетками сетчатой зоны. Клетки пучковой зоны могут быть резервом, активируемым в результате продолжительной стимуляции.
Нормальная функция. Клетки коры надпочечников продуцируют три типа стероидных гормонов.
Минералокортикоиды, в основном альдостерон, синтезируются в клубочковой зоне надпочечников в ответ на ангиотензин II и, в меньшей степени, в ответ на АКТГ. Альдостерон контролирует водно-солевой баланс в основном за счёт стимуляции реабсорбции Na+ в дистальных извитых канальцах почки, а также в слюнных и потовых железах, слизистой оболочке желудка.
Глюкокортикоиды, в основном кортизол и кортикостерон, синтезируются паренхиматозными клетками пучковой зоны и, в меньшей степени, клетками сетчатой зоны, в ответ на АКТГ.
Глюкокортикоиды контролируют метаболизм углеводов, стимулируя как выработку глюкозы из аминокислот и жирных кислот (глюконеогенез), так и превращение глюкозы в гликоген. В коже, мышцах и жировой ткани глюкокортикоиды снижают синтетическую активность и обеспечивают деградацию белков и жиров. Продукты их расщепления, аминокислоты и жирные кислоты используются печенью для глюконеогенеза и синтеза белков. Глюкокортикоиды способны также подавлять иммунный ответ, снижая количество циркулирующих лимфоцитов и эозинофилов.
Андрогены, в основном дигидроэпиандростерон, секретируются в ответ на АКТГ клетками сетчатой зоны и, в меньшей степени, клетками пучковой зоны. Надпочечниковые андрогены имеют менее выраженный маскулинизирующий и анаболический эффект, чем тестостерон.
Функция при патологии
Гиперсекреция. Синдром Кушинга вызван гиперсекрецией кортизола и, зачастую, андрогенов. Клиническими проявлениями этого синдрома являются ожирение, «лунообразное» лицо, гипергликемия, диабет, гирсутизм, аменорея, акне и эмоциональная нестабильность. Гиперсекреция альдостерона (синдром Конна) вызывает задержку Na+ и воды в организме и приводит к повышению артериального давления (гипертензии).
Гипосекреция. Хроническая гипофункция коры надпочечников (Аддисонова болезнь) приводит к снижению уровня глюкозы, натрия, хлорида, бикорбоната в сыворотке крови и повышению уровня калия. В результате больные жалуются на слабость, тошноту, потерю веса и гиперпигментацию. Снижение уровня надпочечниковых андрогенов у женщин при отсутствии компенсации за счёт андрогенов, синтезируемых в яичках, может приводить к снижению оволосения лобка и подмышек.
Фетальная кора располагается между мозговым веществом и тонким слоем неразвитой «постоянной» коры до рождения. Клетки фетальной коры продуцируют сульфатированные андрогены, которые активируются плацентой и попадают в кровоток матери. После рождения клетки фетальной коры дегенерируют, а клетки «постоянной» коры развиваются и образуют три зоны, описанные ранее.
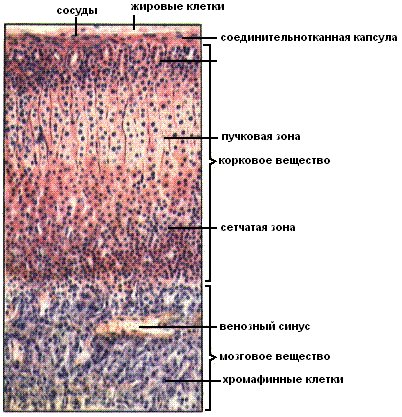
Б. Мозговое вещество надпочечников
Эмбриональное происхождение. Клетки мозгового вещества надпочечников происходят из нервного гребня.
Строение. Состоит из двух основных типов клеток: хромаффинных и ганглионарных.
Хромаффинные клетки (феохромоциты) преобладают среди клеток мозгового вещества, являются постганглионарными симпатическими нейронами, которые утратили аксоны и дендриты. Клетки содержат крупное ядро, большое количество электронно-плотных секреторных гранул заполненных катехоламинами (эпинефрином или норэпинефрином), развитый комплекс Гольджи, небольшое количество ГрЭПС и множество овальных митохондрий. Секреторные гранулы высокочувствительны к хромсодержащим красителям. Хромаффинные клетки синтезируют и выделяют катехоламины в ответ на нервные импульсы, поступающие от преганглионарных симпатических нейронов, расположенных в ядрах боковых рогов спинного мозга (в особенности в ответ на стресс).
Ганглионарные клетки. Небольшое количество парасимпатических ганглионарных клеток имеют типичные для автономных нервных узлов морфологические характеристики. Эти клетки более крупные по сравнению с хромаффинными.
Нормальная функция. Хромаффинные клетки продуцируют два вида катехоламинов в ответ на симпатическую стимуляцию (стресс), которые повышают уровень глюкозы в крови, стимулируя гликогенолиз в печени, и увеличивают приток крови к сердцу. Эпинефрин повышает частоту сердечных сокращений и расширяет кровеносные сосуды в органах, которые необходимы для того, чтобы «бежать» или «бороться», например, в сердце и скелетных мышцах, а также расширяет бронхиолы. В органах, которые играют менее важную роль в борьбе со стрессом (кожа, пищеварительный тракт, почки), эпинефрин и норэпинефрин, наоборот, сужают сосуды. Повышая периферическое сопротивление, эти вещества увеличивают давление и объём крови, притекающей к сердцу, мозгу и скелетным мышцам.
Функция при патологии. Опухоли, состоящие из хромаффинных клеток (феохромоцитомы), приводят к гиперсекреции катехоламинов и, как следствие, состоянию постоянной готовности борьбы со стрессом, даже в его отсутствии, особенно к гипертензии. Опухоли, состоящие из ганглионарных клеток (нейробластомы и ганглионейромы), являются более частыми, особенно у детей, их клинические проявления весьма многообразны.
В. Кровоснабжение надпочечников
- Артерии. Три основных артерии кровоснабжают каждый надпочечник: верхняя надпочечниковая артерия – ветвь нижней диафрагмальной артерии; средняя надпочечниковая артерия отходит непосредственно от аорты; нижняя диафрагмальная артерия – ветвь почечной артерии. Все три артерии прободают капсулу органа поотдельности, а затем их ветви анастомозируют между собой и формируют субкапсулярное артериальное сплетение. Это сплетение даёт начало трём группам артерий:
капсулярные артерии;
корковые артерии, которые ветвятся и дают начало корковым капиллярам, располагающимся между секреторными клетками коркового вещества и опорожняющимся в капилляры мозгового вещества;
артерии мозгового вещества, которые прободают корковое вещество, а по достижении мозгового вещества ветвятся с образованием капилляров мозгового вещества.
Капилляры мозгового вещества получают кровь из артерий как коркового, так и мозгового вещества, а затем формируют несколько мозговых вен.
Вены мозгового вещества формируют единую для каждого надпочечника крупную надпочечниковую вену.
Надпочечниковая вена формируется в центре мозгового вещества и несёт кровь в почечную вену или непосредственно в нижнюю полую вену.
ЩИТОВИДНАЯ ЖЕЛЕЗА
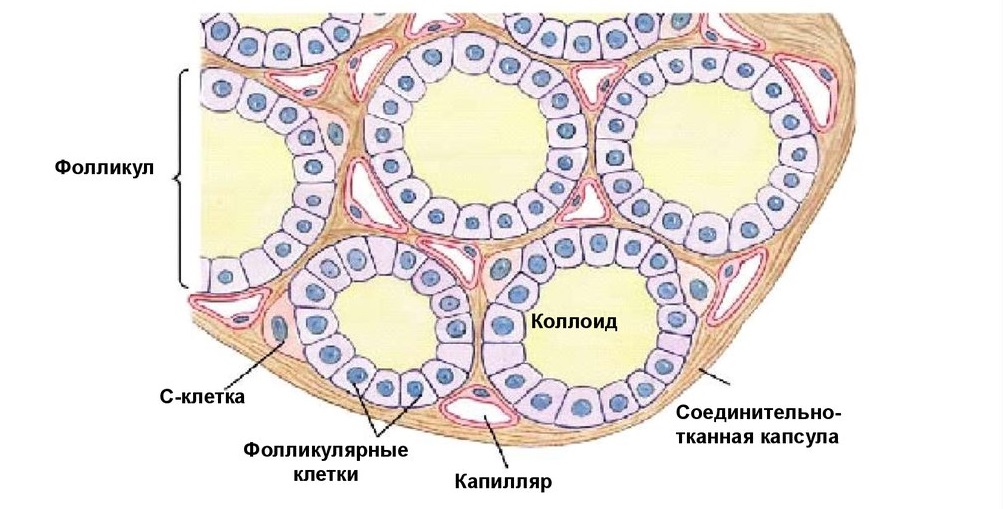
Щитовидная железа начинает развиваться на 4-й неделе эмбриогенеза как выпячивание глоточной энтодермы, и вскоре после этого делится на две доли. У взрослых щитовидная железа располагается в передней области шеи впереди гортани и состоит из двух долей, соединённых перешейком. Каждая доля состоит из множества округлых фолликулов и покрыта тонкой капсулой, от которой вглубь железы отходят септы.
А. Фолликулы щитовидной железы. Каждый фолликул представляет собой шар, стенка которого образована одним слоем эпителиальных фолликулярных клеток, а полость заполнена коллоидом. Фолликулы имеют разный размер и увеличиваются при стимуляции.
Б. Фолликулярные клетки щитовидной железы
Строение. Фолликулярные клетки имеют энтодермальное происхождение и строение, типичное для клеток, продуцирующих пептидные гормоны. Клетки могут быть как плоскими (в неактивном состоянии), так и цилиндрическими при стимуляции.
Нормальная функция. Фолликулярные клетки щитовидной железы отличаются от других эндокринных клеток тем, что накапливают в больших количествах продукт своей секреции (тироглобулин) не в цитоплазматических гранулах, а вне клеток, в коллоиде. Активация деятельности железы происходит при повышенной потребности в энергии под воздействием тиреотропного гормона гипофиза (ТТГ).
Синтез и накопление тироглобулина происходит в несколько этапов: (1) синтез богатого тирозином тироглобулина на ГрЭС; (2) гликозилирование в эндоплазматическом ретикулуме и комплексе Гольджи; (3) упаковка везикул в комплексе Гольджи; (4) слияние везикул с апикальной мембраной клетки и экзоцитоз тироглобулина в просвет фолликула, в коллоид.
Захват и окисление йодида, циркулирующего в крови, обеспечивается с помощью мембранного транспортного белка, располагающегося в базолатеральной мембране фолликулярных клеток. Йодид доставляется в цитоплазму фолликулярной клетки, где он окисляется тиропероксидазой и транспортируется к апикальной поверхности клетки, а затем в полость фолликула. Захват йодида также стимулируется ТТГ.
Йодирование тироглобулина и образование тиреоидных гормонов. На микроворсинках апикальной поверхности фолликулярных клеток, обращенной в коллоид, происходит йодирование тирозиновых остатков тироглобулина. Одна молекула йодида, присоединяясь к тирозину, образует монойодтиронин. Вторая молекула йодида, присоединяясь последовательно к тирозину, даёт начало дийодтиронину. Слияние двух молекул дийодтиронина образует тетрайодтиронин (тироксин, или Т4), тогда как одна молекула монойодтиронина, соединяясь с молекулой дийодтиронина, образует трийодтиронин (Т3). Т4 составляет 90% продуцируемых тиреоидных гормонов, однако он менее активен, чем Т3.
Секреция тиреоидных гормонов. ТТГ стимулирует пиноцитоз коллоида фолликулярными клетками с формированием везикул, содержащих йодированный тироглобулин. Эти везикулы внутри клетки сливаются с лизосомами, содержащими ферменты, которые гидролизируют тироглобулин с образованием тироксина и трийодтиронина, которые выделяются в цитоплазму. Таким образом, происходит высвобождение Т3 и Т4, которые через базальную мембрану проникают в кровоток.
Мишени и эффекты тиреоидных гормонов. Т3 и Т4 ускоряют метаболизм (скорость, с которой клетки потребляют глюкозу), стимулируют рост клеток, ускоряют сердечные сокращения, повышают температуру тела, в общем усиливают все функции клеток, требующие потребления энергии. Также они влияют на клетки гипоталамуса, секретирующие тиролиберин, и тиреотропные клетки аденогипофиза, снижая уровень секреции ТТГ (механизм отрицательной обратной связи).
Функция при патологии
Гипертиреоз – избыточная продукция тиреоидных гормонов (тиреотоксикоз) приводит к нервозности, сердцебиению, учащенному пульсу, мышечной слабости, тремору (дрожанию) конечностей, повышенной утомляемости, потере веса при нормальном аппетите, потливости, постоянному ощущению жары и эмоциональной нестабильности. Увеличивается размер фолликулов за счёт увеличения размеров фолликулярных клеток и увеличения количества накапливаемого тироглобулина, что приводит к гипертрофии самой щитовидной железы – состоянию, известному как зоб.Наиболее известен диффузный токсический зоб (Базедова болезнь, Болезнь Грейвса) – аутоиммунное заболевание, при котором аутоантитела связываются с рецепторами ТТГ на поверхности фолликулярных клеток и постоянно стимулируют их к продукции тироксина.
Гипотиреоз, известный как кретинизм у детей и микседема у взрослых, приводит к снижению потребления глюкозы клетками. Симптомы включают: сонливость, постоянное ощущение холода, замедление мыслительных процессов и моторики, накопление гликозаминогликанов в дерме кожи и, иногда, прибавку в весе.
Так как йодид необходим для нормальной функции щитовидной железы, пониженное содержание йода в пище и воде приводит к функциональному снижению продукции тироксина и часто лежит в основе развития кретинизма и микседемы. Нейодированный тироксин, возникший вследствие дефицита йодида, не обеспечивает отрицательную обратную связь, поэтому гипертрофия фолликулов и зоб могут сопровождать и гипотиреоз.
Болезнь Хашимото – аутоиммунное заболевание, при котором образуются аутоантитела к тироглобулину или тиропероксидазе, что приводит к инфильтрации железы Т-лимфоцитами с последующим разрушением фолликулов.
В. Парафолликулярные клетки (С клетки) расположены диффузно между фолликулярными клетками или образуют скопления между фолликулами. Цитоплазма плохо окрашивается стандартными красителями, поэтому клетки на микропрепаратах выглядят бледными. При электронной микроскопии можно выявить большое количество мелких секреторных гранул. С клетки выделяют пептидный гормон кальцитонин в ответ на повышение уровня кальция в крови. Кальцитонин снижает уровень кальция в крови, стимулируя захват кальция клетками и отложение его в костной ткани.
ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Четыре мелких железы расположены на задней поверхности каждой доли щитовидной железы, по одной у верхних и нижних полюсов. Околощитовидные железы развиваются из энтодермы третьего и четвертого глоточных карманов. Каждая железа покрыта соединительнотканной капсулой, от которой вглубь железы отходят септы, поддерживающие скопления секреторных клеток. У взрослых людей железа состоит из двух основных типов паренхиматозных клеток:
главных и оксифильных.
А. Главные клетки являются самыми многочисленными среди паренхиматозных клеток.
Строение. Мелкие клетки полигональной формы, имеют типичное для клеток, секретирующих пептидные гормоны, строение и множественные мелкие секреторные гранулы, расположенные в бледной цитоплазме.
Нормальная функция. Главные клетки секретируют паратгормон (ПТГ) в ответ на снижение уровня кальция в крови. ПТГ – это пептидный гормон, который воздействует на три органа-мишени для того, чтобы повысить содержание кальция в крови. ПТГ стимулирует резорбцию костной ткани остеокластами. В почках он увеличивает реабсорбцию кальция и выведение фосфатов. Паратгормон также необходим для активации витамина D, который обеспечивает всасывание кальция в кишечнике. Повышение уровня кальция в крови снижает секрецию ПТГ паращитовидными железами.
Функция при патологии
Гиперпаратиреоз. Избыточная секреция ПТГ повышает содержание кальция в крови (гиперкальциемия) и снижает уровень фосфата (гипофасфатемия). Это ведёт за собой повышение уровня кальция в моче, отложению кальция в сосудах и почках и потерю кальция костной тканью, что приводит к остеомаляции и фиброзно-кистозному остеиту.
Гипопаратиреоз. Недостаточная секреция ПТГ нарушает нервно-мышечную проводимость. Низкий уровень кальция в крови вызывает спастические сокращения скелетных мышц и генерализованные судороги, известные как тетания.
Б. Оксифильные клетки – крупные малочисленные клетки паращитовидных желёз, которые за счёт большого количества митохондрий хорошо окрашиваются кислыми красителями. Функция оксифильных клеток неизвестна.
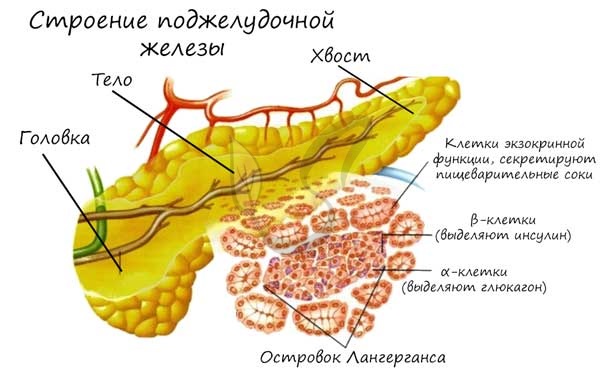
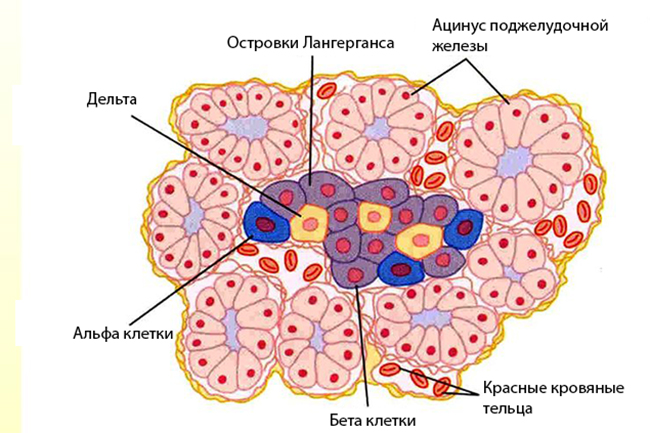
ОСТРОВКИ ЛАНГЕРГАНСА ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
Эти небольшие скопления эндокринных клеток расположены в поджелудочной железе и содержат 4 типа эндокринных клеток.
А-клетки (альфа-клетки). В ответ на низкий уровень глюкозы в крови эти клетки секретируют пептидный гормон глюкагон, эффекты которого противоположны действию инсулина и направлены на повышение уровня глюкозы в крови: в печени расщепляется гликоген (процесс называется гликогенолиз), стимулируется синтез глюкозы из неуглеводных источников (глюконеогенез) – белков и жиров.
Глюкагон также вызывает увеличение секреции инсулина из здоровых β-клеток поджелудочной железы и торможение активности инсулиназы. Это является, по-видимому, одним из физиологических механизмов противодействия вызываемой глюкагоном гипергликемии.
В-клетки (бета-клетки) – наиболее многочисленные клетки островков, секретируют пептидный гормон инсулин в ответ на высокий уровень глюкозы в крови. Синтезированный инсулин сохраняется в клетках в цитоплазматических гранулах в комплексе с цинком. Инсулин связывается с рецепторами на поверхности клеток, что служит сигналом для клеток захватывать глюкозу. В печени под действием инсулина из избытков глюкозы гепатоциты синтезируют гликоген, адипоциты синтезируют триглицериды.
Недостаток инсулина вызывает сахарный диабет I типа, который манифестирует повышением глюкозы в крови (гипергликемия), а затем в моче (глюкозурия). Недостаток или нарушение чувствительности клеточных рецепторов инсулина приводит к развитию сахарного диабета II типа.
Гиперплазия или опухоль из В-клеток может приводить к гиперинсулярному синдрому, который характеризуется гипогликемией.
D-клетки (дельта-клетки) вырабатывают соматостатин, который подавляет секрецию инсулина, глюкагона и гормона роста. Также они могут секретировать гастрин, стимулирующий секрецию желез в слизистой желудка.
Синдром Золингера-Элисона (гастринома) вызывает язву слизистой оболочки за счет гиперсекреции соляной кислоты париетальными клетками желудка. Соматостатинома – редкая опухоль со сложными проявлениями.
F-клетки (или РР-клетки) секретируют панкреатический полипептид, который подавляет экзокринную секрецию ферментов и бикарбоната поджелудочной железой. Также он вызывает расслабление желчного пузыря и снижение секреции желчи.